|
Mimikra chemiczna u mszyc
Dużo pisałem o mimikrze morfologicznej u zwierząt: wyewoluowanym podobieństwie wyglądu jednego gatunku do innego albo do tła środowiskowego. Ta mimikra może chronić zwierzęta przed wzrokiem drapieżników lub, jeśli jesteś drapieżnikiem, ukryć cię przed twoją potencjalną zdobyczą.
Ta druga sytuacja, w której zwierzęta przypominają kogoś lub coś innego, żeby mogły zabić lub zranić członków innych gatunków, nazywa się mimikrą agresywną. O intrygującym nowym przykładzie tego informuje praca w The Proceedings of the National Academy of Sciences autorstwa Adriána Salazara i sześciu kolegów (wolny dostęp, odnośnik poniżej). Ta mimikra jednak dotyczy chemiizamiast wyglądu. W tym nowym przypadku mszyce wyewoluowały do wydzielania węglowodorów, które znajdują się na larwach mrówek, z którymi te mszyce są normalnie stowarzyszone. Ta mimikra oszukuje mrówki, które nie widzą zbyt dobrze, ale są czułe na sygnały chemiczne. Mrówki niosą więc mszyce do mrowiska i wkładają je do komory lęgowej, po czym podstępne mszyce przekłuwają larwy i wysysają hemolimfę.
Cała historia jest bardzo skomplikowana, więc sporo pominę. Mszyca, Paracletus cimiformis, znajduje się na ogół w Europie, ale także w Azji i Afryce Północnej. Żyje na korzeniach roślin i, żeby sprawy jeszcze bardziej skomplikować, jest stowarzyszona z kilkoma gatunkami mrówek. Autorzy badali Tetramorium semilaeve, która jest powszechna w Hiszpanii, gdzie odbywało się badanie. Cykl życia tej mszycy, jak u większości mszyc, jest bardzo złożony, ponieważ żyje naprzemiennie na dwóch roślinach-gospodarzach: drzewie pistacji terpentynowej i na trawach. Dodatkowo mszyce mogą istnieć albo jako uskrzydlone, albo bezskrzydłe oraz jako seksualne i aseksualne. Aseksualne rozmnażają się przez partenogenezę: matka trzyma niezapłodnione, ale diploidalne jaja (stworzone bez normalnego mejotycznego podziału komórki) w swoim ciele i rodzi żywe mszyce. Potomstwo jest więc klonami matki.
Wreszcie – co najważniejsze dla naszych rozważań – mszyce mają dwie interesujące cechy. Pierwszą jest, że dorosłe mszyce żyjące na trawach (ich „wtórnym” gospodarzu) pojawiają się w dwóch postaciach: płaskiej białawej (będę je nazywać „płaskie”) i bardziej zaokrąglonej oliwkowozielonej (będę je nazywał „okrągłe”). Te różnice wyraźnie nie są genetyczne: białe mszyce mogą mieć zielone potomstwo i odwrotnie, prawdopodobnie w zależności od warunków środowiskowych doświadczanych przez rodzica i potomstwo. (Proszę jednak zauważyć, że program pozwalający na taką zamianę jest niewątpliwie zakodowany w genach mszycy.)
Te dwie postaci robią różne rzeczy. Okrągła zielona mszyca jest mutualistą z mrówką: jest trofibiotyczna, co znaczy w tym wypadku, że mszyce, po wyssaniu soku rośliny wydzielają spadź, którą zjadają mrówki. W zamian mrówki dostarczają mszycom ochrony przed drapieżnikami. Jest to więc mutualizm, zachowanie pary gatunków, w którym oba czerpią korzyści ze związku (słynnym przykładem są porosty, które są mieszanką wzajemnie pomagających sobie glonów i grzybów).
Płaska mszyca chemicznie naśladuje larwy mrówek, mrówki zanoszą ją do mrowiska, gdzie wysysa hemolimfę („krew” owadów) z larw mrówek. (Nie jest jeszcze jasne, czy zabija larwy, czy też zmusza je do odrobiny krwiodawstwa, ale badacze znaleźli DNA mrówki w płaskich mszycach i widzieli, jak atakują larwy.) To uważane jest za mimikrę agresywną i można to uważać albo za pasożytnictwo, albo drapieżne żerowanie na mrówkach. A więc mszyce mają skomplikowaną mieszankę postaci uskrzydlonych i bezskrzydłych, rozmnażania płciowego i bezpłciowego, żyją albo na drzewach, albo w trawach i tworzą albo okrągłe postaci mutualistyczne, albo drapieżne białe. (Te różne postaci zwane są „morfami”.) Wykres poniżej pokazuje złożoność; dodałem także podpis z artykułu (nie spolszczony, MK). Możecie to wszystko zignorować poza postaciami okrągłymi i płaskimi po prawej stronie:
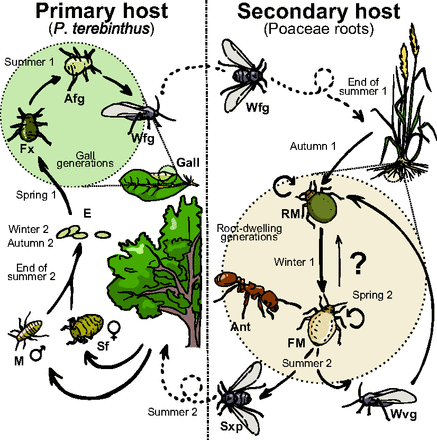
Fig. 1. Simplified diagram of the biannual life cycle of P. cimiciformis. Sexual reproduction takes place on P. terebinthus trees, its primary host, where up to five different morphs occur. Of these generations, three develop inside distinct galls that they induce in their host’s leaves. Toward the end of summer, the last generation born inside the galls consists of winged aphids that fly to the roots of several gramineous species, its secondary host. There, they initiate a succession of root-dwelling wingless parthenogenetic generations consisting of two morphs: the round (RM) and the flat (FM) morphs, respectively. These two morphs participate in mutualistic associations with ants of the genus Tetramorium. In summer, two winged morphs may appear. One disperses the clone to new grasses whereas, in regions where P. terebinthus is present, a second winged morph will fly back to the primary host to give birth to the sexual morphs. Afg, apterous fundatrigeniae; E, eggs; Fx, fundatrix; M, male; Sf, sexual female; Sxp, sexuparae; Wfg, winged fundatrigeniae; Wvg, winged virginoparae. The question mark denotes unclear phenology details during the root-dwelling phase (SI Text and Fig. S1).
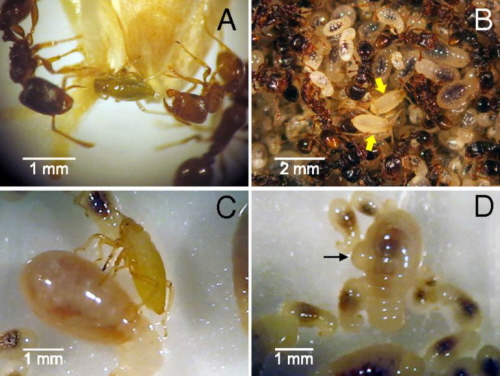
Poniżej jest zdjęcie mrówek i mszyc. Zdjęcie A pokazuje mrówki, które machają czułkami do zielonych, okrągłych mszyc, co jest dla mszyc sygnałem, że mają wydzielić spadź dla mrówek. Obserwacje pokazują, że mrówki machają tylko do okrągłych mszyc, po czym mrówki stukają czułkami w odwłok mszycy, kiedy ta wydziela spadź. Nigdy nie machają do płaskich mszyc, a tylko stukają w nie, które to zachowanie nazywa się „antenacja”.
Zdjęcie B pokazuje płaskie, białe mszyce w komorze lęgowej wśród larw mrówek (strzałki pokazują mszyce, które przypominają larwy). Zdjęcie C pokazuje białą mszycę zatapiającą żuwaczkę w larwie i gotową do wyssania hemolimfy. Zdjęcie D pokazuje larwę, z której wycieka limfa (bąbel) po zaatakowaniu jej:
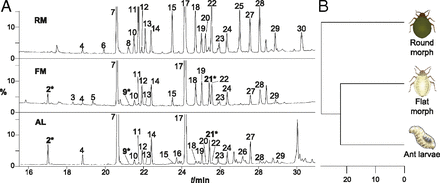
Autorzy sprawdzali hipotezę, że płaskie mszyce mają w sobie coś poza wyglądem, co powoduje, że mrówki biorą je za swoje larwy (nie był to wygląd, bo mszyce płaskie i okrągłe są traktowane zupełnie inaczej także w kompletnej ciemności). Oczywistą hipotezą była „mimikra chemiczna”. Pokazali więc najpierw, używając chromatografii gazowej, że profil węglowodorowy płaskich mszyc jest bardziej podobny do larw mrówek niż mszyc okrągłych.
Praktycznie wszystkie owady mają warstwę węglowodorów na kutykuli; normalnie zapobiegają one wyschnięciu oraz funkcjonują jako sygnały chemiczne. Przez wiele lat pracowałem nad tym u Drosophila i pokazałem, że różne gatunki rozróżniają się wzajemnie, kiedy samce przed kopulacją „kosztują” – używając receptorów chemicznych na swoich przednich odnóżach – węglowodorów znajdujących się na samicach. Można zmienić to rozróżnianie przez przeniesienie węglowodorów od samic innego gatunku, a jak odkryłem, można to zrobić po prostu przez umieszczenie samicy jednego gatunku z mnóstwem samic innego gatunku. Ta samica otrzymuje wiele obcych węglowodorów przez ocieranie się o ciała innych, co wpływa na sposób zalecania się do niej samców.
Najpierw mamy tu profile węglowodorowe (odczyt z chromatografii gazowej) dwóch postaci mszyc i larwy mrówki. Analiza składu (kutykula zawiera wiele węglowodorów) pokazuje, że płaskie mszyce są bardziej podobne do larwy mrówki niż do okrągłej mszycy. Różnice między obydwiema postaciami mszycy nie wynikają po prostu z tego, że nabyła węglowodory od larw przez kontakt z nimi, bo istnieją one również, kiedy mszyce hodowane są w laboratorium bez żadnego kontaktu z mrówkami.
Następnie autorzy sprawdzili, czy różnica węglowodorów ma jakieś znaczenie dla zachowania mrówek. Ma. Ten eksperyment zrobiono przez nasycenie atrap mszyc ekstraktem z larwy mrówki, płaskiej mszycy i okrągłej mszycy oraz rozpuszczalnika heksanu stosowanego jako rozpuszczalnik węglowodorów (grupa kontrolna). Mrówki nie tylko stukały częściej w atrapy z ekstraktem larwy lub płaskiej mszycy niż w atrapy z ekstraktem okrągłej mszycy, ale machały czułkami wyłącznie do atrap z ekstraktem okrągłej mszycy. Ponadto zabierały do mrowiska tylko atrapy z ekstraktem mrówczej larwy lub płaskiej mszycy; nigdy się to nie zdarzyło z atrapą z grupy kontrolnej lub atrapą z ekstraktem z okrągłej mszycy (Jest tam kilka problemów z istotnością statystyczną tych zachowań, więc wyniki są bardziej sugerujące, niż rozstrzygające.)
Profile chemiczne, jak również zachowanie mrówek, sugerują, że płaskie mszyce wyewoluowały do oszukiwania mrówek. Wydaje się to być stanem pochodnym, bo wszystkie pokrewne mszyce, jakie znamy, mają tylko postać mutualistyczną i okrągłą, wydzielającą spadź.
Wywołuje to kilka pytań. Poruszę tylko dwa. Po pierwsze, jak to wyewoluowało? Chociaż postaci płaska i okrągła są podobne genetycznie, a różnice między nimi kontrolowane są przez jakieś sygnały środowiskowe (jakie sygnały jest kolejnym pytaniem), program genetyczny, który powoduje, że mszyca jest albo okrągła, albo płaska, znajduje się w genomie mszycy; jest to program kierowany w jednym lub drugim kierunku przez sygnały środowiskowe. Program i czułe na środowisko przełączniki są z pewnością produktem doboru naturalnego. Jak jednak płaska postać wyewoluowała z okrągłej, choć obie współistnieją, jest zagadką; nie wiemy także zbyt dużo o innych aspektach ewolucji skomplikowanego cyklu życiowego mszyc. W tej mszycy jest wiele programów genetycznych (okrągłe kontra płaskie, seksualne kontra aseksualne, uskrzydlone kontra bezskrzydłe, mieszkające na drzewach kontra mieszkające na trawach) i o zawrót głowy przyprawia rozważanie, jak mogły wyewoluować. (Przypuszczam, że Discovery Institute użyje naszej niewiedzy, żeby zakrzyknąć: „Bóg to zaprojektował!”)
Po drugie, co utrzymuje obie postaci mszyc w jednej populacji? Jedną oczywistą odpowiedzią jest rodzaj „doboru zależnego od częstotliwości występowania”. To jest, chociaż różnice między mszycami płaskimi a okrągłymi oparte są na sygnałach środowiskowych, program genetyczny wyewoluował prawdopodobnie, by reagować na te sygnały w sposób adaptacyjny, dając różne postaci, kiedy są one najbardziej adaptacyjne. Jedną - moją - teorią jest to, że kiedy zaczyna być zbyt dużo płaskich mszyc, okrągłe mają przewagę, bo liczebność potomstwa mrówek będzie tak drastycznie zmniejszona, że kolonia może wymrzeć, zagrażając przetrwaniu wszystkich mszyc stowarzyszonych z danym mrowiskiem.
I odwrotnie, kiedy jest bardzo dużo okrągłych mszyc, korzystne może być unikanie konkurencji przez tworzenie postaci płaskich, które zajmują zupełnie inną niszę pokarmową.
Te korzyści zależą od stosunkowej częstości występowania tych dwóch postaci i dlatego nazywa się to doborem zależnym od częstości występowania. (Jest to dobór, który działa nie na programie genetycznym kodującym dwa różne wyglądy i zachowania, ale na programie genetycznym do ustalania, kiedy mszyce zmieniają się z jednej postaci w drugą.)
Wreszcie można też rozważać różnice strategii ewolucyjnej tych dwóch postaci, jedna mutualistyczna, a druga pasożytnicza. Tutaj jednak zacytuję po prostu słowa autorów:
Dwoista strategia mszyc P. cimiciformis przedstawia skomplikowany scenariusz ewolucyjny. Z jednej strony, okrągły morf i mrówki zaangażowane w stosunek trofibiotyczny powinny być przedmiotem konfliktu interesów typowego dla mutualizmu, z doborem popychającym każdego partnera do maksymalizowania własnych korzyści przez dawanie minimum własnej energii i zasobów. Z drugiej strony, płaski morf i mrówki mogą być zaangażowane w wyścig zbrojeń, z doborem faworyzującym coraz lepsze zdolności oszukiwania mrówek przez mszyce i coraz lepsze umiejętności wykrywania nienależących do kolonii członków przez mrówki.
Nawiasem mówiąc, jeśli zajrzycie na stronę tej pracy, możecie obejrzeć trzy dodatkowe filmy o zachowaniu mrówek/mszyc i o ich interakcjach, które nie pojawiają się w artykule.
________________
Salazar, A. et al. 2015. Aggressive mimicry coexists with mutualism in an aphid. Proceedings of the National Academy of Sciences 112 1101-1106; published ahead of print January 12, 2015, doi:10.1073/pnas.1414061112 Chemical mimicry in an aphid Why Evolution Is True, 28 stycznia 2015 Tłumaczenie: Małgorzata Koraszewska Jerry A. Coyne
Profesor na wydziale ekologii i ewolucji University of Chicago, jego książka "Why Evolution is True" (Polskie wydanie: "Ewolucja jest faktem", Prószyński i Ska, 2009r.) została przełożona na kilkanaście języków, a przez Richarda Dawkinsa jest oceniana jako najlepsza książka o ewolucji. Jerry Coyne jest jednym z najlepszych na świecie specjalistów od specjacji, rozdzielania się gatunków. Jest wielkim miłośnikiem kotów i osobistym przyjacielem redaktor naczelnej.
Znalezionych 1476 artykuły.
|
|
|
|
|
|
|
| Dlaczego kod genetyczny nie jest uniwersalny |
|
Cobb |
|
2014-10-06 |
| Zachwycający rabuś, który liczy sto milionów lat |
|
Cobb |
|
2014-04-28 |
| Twoja wewnętrzna mucha |
|
Cobb |
|
2014-01-22 |
| Ćma gynandromorf wychodzi na światło dzienne - opowiada historię o nauce |
|
Cobb |
|
2015-09-15 |
| Seksizm w nauce: czy Watson i Crick naprawdę ukradli dane Rosalind Franklin? |
|
Cobb |
|
2015-07-07 |
| Uroczy wykres, który opowiada naszą historię |
|
Cobb |
|
2017-10-17 |
| 12 podstawowych punktów biologii ewolucyjnej |
|
Cobb |
|
2016-03-02 |
| Świat RNA |
|
Cobb |
|
2014-11-27 |
| Jak karakara wygrywa z osami |
|
Cobb |
|
2013-12-29 |
| Seymour Benzer: humor, historia i genetyka |
|
Cobb |
|
2014-03-21 |
| Dlaczego powinny nas fascynować liczące 100 tysięcy lat ludzkie zęby z Chin? |
|
Cobb |
|
2015-10-30 |
| DNA: zoptymalizowany kod źródłowy? |
|
Cobb |
|
2015-11-30 |
| Urodziny Rosalind Franklin! |
|
Cobb |
|
2020-07-31 |
| Wszystkiego najlepszego w dniu 60. urodzin, centralny dogmacie! |
|
Cobb |
|
2017-10-04 |
| Dziwaczne, wysysające krew czerwie jurajskie |
|
Cobb |
|
2014-06-28 |
| Geny neandertalskie są wszędzie |
|
Cobb |
|
2015-10-23 |
| Technologia pomaga w kryzysach wodnych na całym globie |
|
Cohen |
|
2019-04-02 |
| Ptasia grypa w czasach ludzkiej zarazy |
|
Collins |
|
2022-01-11 |
| Oszaleć na punkcie nietoperzy w czasach korony i politykierstwa |
|
Collins |
|
2020-07-25 |
| Oxitec rozszerza próby z komarami GMO, by zredukować szerzenie się malarii |
|
Conrow |
|
2022-04-28 |
| Nigeria daje zielone światło kukurydzy GMO |
|
Conrow |
|
2021-11-22 |
| Rośliny zmodyfikowane: odkłamać opinię o GMO |
|
Conrow |
|
2022-04-07 |
| Bakłażan GMO jest udokumentowaną wygraną ubogich farmerów |
|
Conrow |
|
2021-09-23 |
| Selektywnie stosowana koncepcja tabula rasa i ideologicznie motywowane nieporozumienia |
|
Cory Clark |
|
2019-05-09 |
| Dlaczego zwierzęta są urocze? |
|
Coyne |
|
2014-12-30 |
| Trzecia droga ewolucji? Nie sądzę |
|
Coyne |
|
2015-03-05 |
| Lekcja ewolucji: specjacja w akcji! |
|
Coyne |
|
2015-01-12 |
| Moda na kopanie nauki |
|
Coyne |
|
2014-02-03 |
| Niezwykłe pasikoniki naśladujące liście, u których samce i samice są różnych kolorów |
|
Coyne |
|
2017-01-24 |
| Francis Crick był niesamowitym geniuszem |
|
Coyne |
|
2015-04-02 |
| Ogon ćmy i nietoperze |
|
Coyne |
|
2015-02-23 |
| Skąd bóbr? To są szczuroskoczki, a nie wiewiórki! |
|
Coyne |
|
2017-04-11 |
| Dan Brown - akomodacjonista |
|
Coyne |
|
2015-01-31 |
| Ideologiczna opozycja wobec prawdy biologicznej |
|
Coyne |
|
2016-12-28 |
| Nowe niezwykłe skamieniałości typu “Łupki z Burgess” |
|
Coyne |
|
2014-02-22 |
| Kolejny gatunek wron używa narzędzi |
|
Coyne |
|
2016-10-06 |
| Seks paproci i kreacjoniści |
|
Coyne |
|
2015-03-27 |
| Mistyfikacja Sokala: dwadzieścia lat później |
|
Coyne |
|
2017-01-13 |
| Dobór naturalny w naszym gatunku na przestrzeni ostatnich dwóch tysiącleci |
|
Coyne |
|
2016-10-22 |
| Nowa data udomowienia kotów: około 5300 lat temu – i to w Chinach |
|
Coyne |
|
2013-12-26 |
| Pisklę przypominające wyglądem i zachowaniem trującą gąsienicę |
|
Coyne |
|
2014-12-18 |
| Czy rozum jest “większy niż nauka”? Kiepska próba deprecjonowania nauki |
|
Coyne |
|
2015-04-28 |
| Ewolucyjny poziom ludzkiej przemocy |
|
Coyne |
|
2016-10-14 |
| Ciąg dalszy sporu o dobór grupowy |
|
Coyne |
|
2015-04-22 |
| Eureka! Sprytne wrony to odkryły |
|
Coyne |
|
2014-04-07 |
| Koniec humanistyki? |
|
Coyne |
|
2014-01-15 |
| Nowe skamieniałości: najwcześniejszy na świecie znany ptak |
|
Coyne |
|
2015-05-12 |
| Facet od nauki przeciwko GMO |
|
Coyne |
|
2014-11-12 |
| Najstarsze organizmy: 3,7 miliarda lat? |
|
Coyne |
|
2016-09-13 |
| Montezuma i jego flirty |
|
Coyne |
|
2014-05-11 |
| Specjacja hybryd może być rzadka |
|
Coyne |
|
2016-10-29 |
| Trawa w uchu. Ale po co? |
|
Coyne |
|
2014-07-09 |
| Koszmar kreacjonisty: ewolucja w działaniu |
|
Coyne |
|
2016-09-21 |
| Nowy, opierzony i czteroskrzydły dinosaur |
|
Coyne |
|
2014-07-23 |
| Zmień swoje geny przez zmianę swojego życia |
|
Coyne |
|
2014-05-15 |
| Czy człowiek musiał wyewoluować? |
|
Coyne |
|
2015-05-15 |
| Selektywne używanie narzędzi wśród mrówek |
|
Coyne |
|
2017-01-17 |
| Adam i Ewa: dwoje, czy więcej niż dwoje przodków? |
|
Coyne |
|
2017-01-07 |
| Historia porostów i człowieka, który ją skorygował |
|
Coyne |
|
2017-01-26 |
| Delfiny umyślnie narkotyzują się truciznami rozdymków |
|
Coyne |
|
2014-01-04 |
| Przedwczesna wiadomość o śmierci samolubnego genu |
|
Coyne |
|
2013-12-22 |
| Homo floresiensis, hominin “hobbit”, w Internecie |
|
Coyne |
|
2016-11-25 |
| Modliszka storczykowa: czy upodabnia się do storczyka? |
|
Coyne |
|
2015-03-13 |
| Cuda genetyki: arbuz bez pestek |
|
Coyne |
|
2014-08-25 |
| Pająk upodabnia się do ptasich odchodów |
|
Coyne |
|
2014-06-17 |
| Ewolucja i Bóg |
|
Coyne |
|
2014-01-29 |
| Intelektualne danie dnia The Big Think |
|
Coyne |
|
2014-03-04 |
| Tajemnica pasków zebry rozwiązana – a przynajmniej tak mówią naukowcy |
|
Coyne |
|
2017-01-31 |
| Mimikra chemiczna u mszyc |
|
Coyne |
|
2015-02-19 |
| Jak często geny przeskakują między gatunkami? |
|
Coyne |
|
2015-04-18 |
| Marnie napisany artykuł o uroczym gryzoniu |
|
Coyne |
|
2014-07-03 |
| Ślepa salamandra z Teksasu ma nerw wzrokowy, ale nie ma prawdziwych oczu |
|
Coyne |
|
2016-10-11 |
| Ukwiał żyje w antarktycznym lodzie! |
|
Coyne |
|
2014-01-25 |
| Żebropławy, czyli dziwactwa ewolucji |
|
Coyne |
|
2013-12-30 |
| OLBRZYMI owad wodny (i kilka innych) |
|
Coyne |
|
2014-07-28 |
| Seksizm w nauce o jaskiniowych owadach |
|
Coyne |
|
2014-04-26 |
| Dobór krewniaczy pozostaje wartościowym narzędziem |
|
Coyne |
|
2015-04-06 |
| Najstarsza jak dotąd identyfikacja medycyny sądowej |
|
Coyne |
|
2014-12-10 |
| Pradawnym płazom odrastały kończyny |
|
Coyne |
|
2014-09-29 |
| Dymorfizm płciowy i ideologia |
|
Coyne |
|
2014-12-01 |
| Bajka o kaczkach karolinkach |
|
Coyne |
|
2016-12-16 |
| Grantowie na Galápagos i ich hybrydowe gatunki |
|
Coyne |
|
2014-08-18 |
| Czy humaniści boją się nauki? |
|
Coyne |
|
2014-02-07 |
| Fantastyczna mimikra tropikalnego pnącza |
|
Coyne |
|
2014-05-03 |
| Nowy opierzony dinozaur sugeruje, że większość dinozaurów miała pióra |
|
Coyne |
|
2014-08-03 |
| Dowody ewolucji: wideo i nieco dłuższy wywód |
|
Coyne |
|
2014-10-22 |
| O pochodzeniu dobra i zła |
|
Coyne |
|
2014-02-05 |
| Genetyka kocich łat |
|
Coyne |
|
2014-11-26 |
| Wierzący nagradzani za życia |
|
Coyne |
|
2014-12-21 |
| Lucy mogła umrzeć spadając z drzewa |
|
Coyne |
|
2016-09-07 |
| Opierzony ogon dinozaura w bursztynie! |
|
Coyne |
|
2016-12-19 |
| Nowa i dziwaczna, zmieniająca kształt żaba |
|
Coyne |
|
2015-04-10 |
| Z nowego artykuły wynika, że istnieje nie jeden, a cztery gatunki żyraf, nie jestem jednak pewien |
|
Coyne |
|
2016-09-27 |
| John van Wyhe obala mity o Darwinie |
|
Coyne |
|
2016-11-09 |
| Ewolucja, ptaki i kwiaty |
|
Coyne |
|
2014-06-02 |
| Pradawne rośliny kwitnące znalezione w bursztynie |
|
Coyne |
|
2014-01-10 |
| Najstarsze żyjące organizmy |
|
Coyne |
|
2014-04-03 |
| Użycie ognia przez homininy: przykład szybkiej ewolucji kulturowej? |
|
Coyne |
|
2021-08-04 |
| Cztery prawa biologii ewolucyjnej |
|
Coyne |
|
2015-10-13 |
| Znaleziono najstarszego “bilaterian”: odkryto podobne do robaka stworzenie wraz z jego skamieniałymi śladami |
|
Coyne |
|
2020-04-16 |
|
| « Poprzednia strona Następna strona » |
| | |