|
Moja ostatnia praca badawcza: Część 1: Cele i metody
|
Jerry A. Coyne
| 2020-02-01
|
Moja ostatnia praca “badawcza” – taka, do której zbierałem dane własnymi rękami, popychając muchy (tj. manipulując Drosophila pod mikroskopem stereoskopowym) – została wreszcie opublikowana w „Genetics” i z przyjemnością zobaczyłem, że jest “podkreślonym artykułem”. No cóż, sam także uważam, że praca jest dobra i jest dobrym zakończeniem mojej kariery popychacza muszek. Co więcej, artykuł jest dostępny dla każdego i można przeczytać opublikowaną wersję po kliknięciu na link pod zrzutem z ekranu lub uzyskać pdf tutaj (pełen odnośnik pod spodem). Opisanie całej pracy, włącznie z naszymi celami, metodami i wynikami (a jest ich dużo!) byłoby zbyt długie i nużące. Postanowiłem więc, że zrobię z tego dwa posty, jeden o celach i metodach i drugi o wynikach. Mam nadzieje, że łatwiej to będzie strawić. Tutaj jest część 1. Pisałem pokrótce o tym artykule trzy dni temu, ale tutaj mogę napisać więcej. Wynikł z pracy, jaką niemal każdy genetyk, którego znam, chciał zrobić, ale ten rodzaj badania jest ryzykowny, bo jest eksploracyjne, a co ważniejsze, trwa bardzo długo. Z grantami przyznawanymi na około trzy lata nie ma sposobu na przeprowadzenie tego badania i skończenie go w okresie jednego grantu, a to znaczy, że jeśli zaczynasz badanie finansowane przez grant, nie masz niemal niczego do pokazania, kiedy przychodzi pora odnowienia grantu. Ja napisałem propozycję tego badania jako część mojego ostatniego grantu z NIH, więc nie musiałem martwić się o posiadanie rezultatów po trzech latach.
Bardzo jednak chciałem wykonać to badanie i kiedy mówiłem o wynikach na różnych uniwersytetach, niemal zawsze spotykałem badacza, który mówił: „Wiesz, także myślałem o takim badaniu”. Powodem nie jest jakiś wspaniały pomysł, ale częste pytanie: „Co dzieje się, kiedy dwa gatunki tworzą hybrydy tak, że ich genomy są całkowicie wymieszane?” W takich wypadkach otrzymujemy „hybrydowy rój” i chcemy wiedzieć, co się z nim dzieje. Jest kilka możliwości:
a.) Może pozostać rojem mieszańców z segregującymi genami z obu gatunków.
b.) Hybrydowy rój może pozostać mieszany do pewnego stopnia i wyewoluować w nowy gatunek, który jest reprodukcyjnie izolowany od obu gatunków rodzicielskich. To nazywa się „diploidalnym gatunkiem hybrydowym” i zdarza się w naturze, ale nieczęsto. (Jednak gatunki wynikające z hybrydyzacji, a następnie podwajające liczbę chromosomów, zwane „gatunkami poliploidalnymi”, są dość częste, szczególnie wśród roślin.)
c.) Rój może powrócić do jednego z gatunków rodzicielskich, tracąc przez dobór geny drugiego gatunku. Jeśli tworzysz repliki hybrydowych rojów, możesz zobaczyć czy, jeśli to się zdarza, zawsze powraca do tego samego gatunku rodzicielskiego, czy też w różnych replikach może powrócić do jednego lub drugiego gatunku rodzicielskiego.
Spodziewaliśmy się, że najbardziej prawdopodobnym scenariuszem będzie c), ale nie byliśmy pewni. Powodem, że tak sądziliśmy, jest to, że w obu parach gatunków, jakich użyliśmy, hybrydowe samce były w znacznej mierze sterylne, a więc dobór dokonałby czystki genów powodujących bezpłodność. Nie znaczy to jednak, że wszystkie geny mogłyby zostać wyeliminowane. A nawet jeśli zaszedłby powrót, nie wiedzieliśmy, jak szybko to się stanie ani czy wpłynie w zróżnicowany sposób na widoczne cechy, które rozróżniają te gatunki, na międzygatunkowe zachowania kopulacyjne gatunków, lub na DNA.
Mogliśmy także stworzyć w laboratorium nowy, hybrydowy gatunek! Po prostu nie wiedzieliśmy, co się zdarzy, ale wiedzieliśmy, że coś się zdarzy i że będzie to interesujące.
Stworzyliśmy więc roje hybrydowe. Stworzyliśmy je z dwóch różnych par „siostrzanych gatunków” Drosophila (najbliższymi wzajemnymi krewnymi) i dla każdego roju stworzyliśmy osiem replik. To dało nam 16 możliwości zobaczenia, co się stanie. I w każdej z tych 16 replik (każda składająca się z jednej butli hybryd) śledziliśmy morfologię, zachowanie i sekwencje DNA przez 20 lub więcej pokoleń – około rok w laboratorium – żeby zobaczyć, co się stanie. To był wielki eksperyment, ponieważ mierzyliśmy wiele cech w wielu replikach, badaliśmy zachowanie i płodność samców i sekwencjonowaliśmy całe genomy każdej repliki: ponad 20 milionów nukleotydów.
Podam wam rezultaty w jednym słowie: powrót i to konsekwentny powrót do jednego rodzica. Ale więcej o tym następnym razem.
Poniżej są dwie pary gatunków, składające się na dwie odrębne hybrydyzacje (każdy rój hybryd był tworzony 8 razy niezależnie dla każdej pary). I każdy rój hybrydowy był tworzony przez wzajemne krzyżowanie gatunków, dając dwa typy hybrydowych samic, każda zawierająca połowę DNA każdego gatunku rodzicielskiego; połowa z nich z cytoplazmą i mitochondriami jednego gatunku rodzicielskiego (matki), a połowa samic mająca te cechy od drugiego gatunku. Teraz mieliśmy hybrydy samic do rozpoczęcia roju z równymi proporcjami całego DNA, cytoplazmy i organelli. Ponieważ jednak hybrydy samców były sterylne, potrzebowaliśmy samców, by hybrydowy rój zaczął się rozmnażać; dodaliśmy więc do tych samic równe liczby samców z obu rodzicielskich gatunków (samce stworzone przez to krzyżowanie wsteczne z hybrydowymi samicami są częściowo płodne). Hybrydowe samice nie wykazywały dyskryminacji przy krzyżowaniu się, więc kopulowały z samcami obu gatunków, dając na wpół płodne samce i w pełni płodne samice, a potem pozwoliliśmy rojom rozmnażać się przez 20-24 pokolenia. Oczywiście, płodność poprawiła się w trakcie trwania eksperymentu, w miarę jak geny dające męską sterylność były usuwane przez dobór. Hybrydowe roje zaczęły z czystą 50/50 mieszanką DNA, chromosomami (włącznie z Y) i organellami + cytoplazma z obu gatunków.
Dwie pary gatunków, które skrzyżowaliśmy:
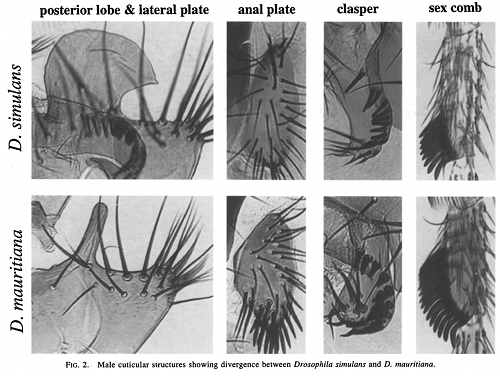
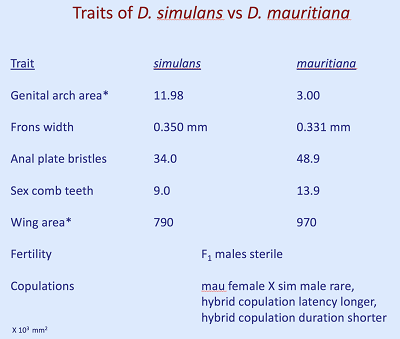
1.) Drosophila simulans/D. mauritiana. D. simulans jest komensalna dla ludzi na całym świecie, podczas gdy D. mauritiana znajduje się tylko na Oceanie Indyjskim na Mauritiusie, gdzie jest endemiczna. Sekwencjonowanie DNA ustala ich rozejście się na między 250 tysięcy a 500 tysięcy lat temu. Choć wyglądają bardzo podobnie, samce różnią się szeregiem drugorzędowych cech płciowych, co sugeruje stosunkowo szybki dobór płciowy (samic nie można rozróżnić, oglądając je). Oto niektóre różnice w kształcie genitalnych walw samca, liczbie szczecinek na odbycie, liczbie szczecinek na genitalnych walwach, liczbie „zębów” w grzebieniu płciowym, strukturze na przednich odnóżach samców, których używają do przytrzymywania samic podczas kopulacji. Najpierw, ilustracje:
A tutaj są przeciętne różnice między gatunkami dla kilku cech (błędy standardowe nie pokazane): Tak więc podczas trwania eksperymentu mogliśmy mierzyć te cechy, jak również inne (szerokość czoła, powierzchnię skrzydła i tak dalej), żeby zobaczyć, czy wartości cech pozostają pośrednie w hybrydach, czy też populacja zaczyna powracać do wartości cech charakterystycznych dla jednego lub drugiego gatunku.
Ponadto, gatunki pokazały znaczącą dyskryminację w krzyżowaniu się: chociaż samice D. simulans chętnie kojarzyły się w samcami D. mauritiana, odwrotne krzyżówki niemal nigdy nie zdarzają się: samice mauritiana po prostu odrzucają natrętnie zalecających się samców simulans. To daje nam możliwość zobaczenia, czy zachowania godowe hybryd powracają do jednego lub do drugiego gatunku. Można to osądzić przez krzyżowanie samców z hybrydowych rojów w różnych okresach z samicami D. mauritiana. Jeśli są odrzuceni, to stają się podobni do simulans w męskich cechach, które samice odrzucają, ale jeśli zostają zaakceptowani, to te cechy powracają do cech mauritiana.
Dalej: kiedy kopulacje jednak zachodzą między samcami simulans a samicami mauritiana, są krótkie i nienormalne: samice nie lubią samców wspinających się na ich grzbiet i próbują ich skopać, więc „kopulacja” obejmuje sytuacje, w których samce są ciągnięte po laboratoryjnej probówce za genitalia. To wygląda na bolesne! Ten rodzaj dziwacznej kopulacji jest nienormalnie krótki i często kończy się bez zapłodnienia. Kopulacja simulans X simulans trwa około 30 minut, kopulacja mauritiana X mauritiana trwa około 13 minut, a nienormalna kopulacja hybryd trwa około ośmiu minut.
To daje nam inny sposób mierzenia cech godowych: użycie samca z hybrydowego roju i umieszczeniu go z samicą mauritiana . Jeśli kopulacja jest nienormalna i trwa około 8 minut, ma on cechy, które powróciły do cech D. simulans. Jeśli kopulacja jest normalna i trwa około 13 minut, samce powróciły do D. mauritiana.
Wreszcie, ponieważ hybrydowe samce między dwoma czystymi gatunkami są sterylne, możemy zobaczyć, czy samce i samice w hybrydowych rojach powróciły do jednego lub drugiego gatunku pod względem genów sterylności. Po prostu skrzyżuj samicę z roju z samcami D. mauritiana i, osobno, z samcami D. simulans. Jeśli produkuje płodne hybrydy z pierwszymi, ale nie z drugimi, powróciła do D. mauritiana. Jeśli otrzymujemy odwrotny rezultat, powróciła do D. simulans. Można to zrobić także dla samców i zobaczyć, czy mają związek sterylności z jednym czystym gatunkiem, czy z drugim.
Na koniec jest DNA. Te gatunki różnią się liczbą miejsc w DNA, a więc można zidentyfikować odcinek genomu z jednego lub drugiego gatunku po prostu przez sekwencjonowanie go. Czy ma sekwencje jak mauritiana, czy jak simulans? Aby to zrobić zsekwencjonowaliśmy całe genomy wszystkich hybrydowych rojów w 20 pokoleniu, żeby zobaczyć, czy ich genomy były nadal bardzo zmieszane, czy też przesunęły się w kierunku jednego lub drugiego gatunku. (Znaliśmy pełne sekwencje DNA dwóch gatunków rodzicielskich.) To była żmudna część eksperymentu, bo musieliśmy najpierw mieć sekwencje czystych gatunków, znaleźć różnice między nimi, a potem porównać te sekwencje z hybrydami.
Tak więc użyliśmy kombinacji cech morfologicznych, behawioralnych, płodności i DNA, żeby dowiedzieć się, co stało się z rojami hybrydowymi. Zrobiliśmy to samo dla innej pary gatunków:
2.) Drosophila yakuba/D. santomea. Spędziłem ostatnie dziesięć lat w laboratorium pracując z tą parą gatunków, bo D. santomea (odkryta przez mojego nieżyjącego kolegę, Daniela Lachaise’a) jest endemiczna dla wyspy São Tomé, około 250 km na zachód od Gabonu. Na tej wyspie jest jednak także jej siostrzany gatunek, D. yakuba, a rozdzieliły się one około milion lat temu. D. yakuba jest rozprzestrzeniona na lądzie afrykańskim, znajduje się na sawannach, łąkach i w lasach. D. santomea prawdopodobnie nie powstała na wyspie w obecności swojego siostrzanego gatunku, ale raczej ich wspólny przodek dokonał inwazji na wyspę milion lat temu. Kiedy przodek wyewoluował przez transformację najeźdźcy w D. santomea na wyspie, D. yakuba najechała ponownie, prawdopodobnie w ciągu ostatnich 10 tysięcy lat.
Te dwa gatunki współistnieją teraz na wyspie, ale yakuba żyje na niższych wysokościach, a santomea jest ograniczona do wysokości powyżej 1000 m. Istnieje strefa hybrydowa między nimi, tam gdzie się stykają, więc mogliśmy zobaczyć, co dzieje się w naturze, kiedy te dwa gatunku hybrydyzują.
W obu wypadkach więc stworzyliśmy repliki między szeroko rozprzestrzenionym „kontynentalnym” gatunkiem (nazywamy go w artykule „dominującym”) a gatunkiem endemicznym dla wyspy.
Podobnie jak simulans/mauritiana, yakuba/santomea także mają różne cechy morfologiczne, choć nie różnią się tak głęboko. Najbardziej uderzająca jest różnica pigmentacji. Podobnie jak 7 innych gatunków w podgrupie D. melanogaster, D. yakuba ma czarną pigmentację zarówno u samców, jak samic, a jest ona szczególnie wyraźna u samców na czubku odwłoka. D. santomea, z drugiej strony, jest wyjątkowa w tym, że niemal całkowicie brak jej czarnego pigmentu, jak widać na poniższym zdjęciu dwóch samców:
To daje nam sposób mierzenia ewolucji hybrydowego roju, bo stworzyliśmy sposób ilościowej oceny czerni na skali od 0 do 120. (Wszystkie muszki były dobierane do oceny na ślepo.)
Yakuba/santomea różnią się także przeciętnie w dwóch morfologicznych cechach: liczbie szczecinek hypandrial (na genitaliach samców) i liczbie „zębów” w płciowych grzebieniach, choć różnica nie jest tak wyraźna jak między simulans i mauritiana. (Ja odkryłem różnicę w liczbie zębów grzebieni płciowych.) Możemy więc mierzyć te trzy cechy wraz z upływem czasu, żeby zobaczyć, jak ewoluuje rój hybrydowy yakuba/santomea.
Stosunki sterylności między tą parą są podobne do tej drugiej pary: męskie hybrydy są sterylne. Możemy więc wziąć samce i samice z hybrydowego roju, skojarzyć je z członkami czystych gatunków i zobaczyć, czy rój przyjmuje geny płodności z jednego gatunku, czy z drugiego.
Czas od momentu, kiedy samiec spotyka się z samicą do chwili kopulacji oraz długość trwania kopulacji także różnią się między gatunkami, dając nam kolejną behawioralną cechę do mierzenia, przez krzyżowanie osobników z hybrydowego roju z osobnikami czystymi gatunkowo.
Wreszcie, gatunki z tej pary, podobnie jak z tej drugiej, różnią się sekwencjami DNA w sposób charakterystyczny dla gatunku, więc mogliśmy sekwencjonować DNA członków hybrydowego roju yakuba/santomea w 20. pokoleniu, żeby zobaczyć, jak wyewoluował. Czy nadal była to wielka mieszanka genomów gatunków rodzicielskich, czy też był powrót do sekwencji jednego lub drugiego gatunku?
W następnym poście (mam nadzieję, że w ciągu kilku dni) opiszę wyniki, koncentrując się na roju mauritiana/simulans, ponieważ wyniki są właściwie takie same dla obu par gatunków i nie ma potrzeby podawać wszystkich wyników, skoro są one w artykule.
Próbowałem streszczać się, ale, jak widzicie, nadal zabiera sporo miejsca opisanie tego, co zrobiliśmy. (I napisanie tego postu zabrało mi dwie godziny!).
________________ Matute, D. R., A. A. Comeault, E. Earley, A. Serrato-Capuchina, D. Peede, A. Monroy-Eklund, W. Huang, C. D. Jones, T. F. C. Mackay, and J. A. Coyne. 2020. Rapid and predictable evolution of admixed populations between two Drosophila species pairs. Genetics 214:211-230. My last research paper: Part1: Aims and methods Why Evolution Is True, 26 stycznia 2020 Tłumaczenie: Małgorzata Koraszewska Jerry A. CoyneEmerytowany profesor na wydziale ekologii i ewolucji University of Chicago, jego książka "Why Evolution is True" (Polskie wydanie: "Ewolucja jest faktem", Prószyński i Ska, 2009r.) została przełożona na kilkanaście języków, a przez Richarda Dawkinsa jest oceniana jako najlepsza książka o ewolucji. Jerry Coyne jest jednym z najlepszych na świecie specjalistów od specjacji, rozdzielania się gatunków. Jest również jednym ze znanych "nowych ateistów" i autorem książki "Faith vs Fakt". Jest wielkim miłośnikiem kotów i osobistym przyjacielem redaktor naczelnej.
Znalezionych 1475 artykuły.
|
|
|
|
|
|
|
| Tajemnice życia płodowego |
|
Zimmer |
|
2014-06-07 |
| Czy ludzkość zmierza w kierunku kanibalizmu? |
|
Lomborg |
|
2014-06-09 |
| Milczenie świerszczy |
|
Yong |
|
2014-06-11 |
| Maccartyzm w klimatologii |
|
Lomborg |
|
2014-06-12 |
| Życie w powiększeniu |
|
Zimmer |
|
2014-06-13 |
| Pół miliarda lat samobójstw |
|
Yong |
|
2014-06-14 |
| Amfisbeny |
|
Naskręcki |
|
2014-06-16 |
| Pająk upodabnia się do ptasich odchodów |
|
Coyne |
|
2014-06-17 |
| Tajemny składnik młodej krwi: oksytocyna? |
|
Zimmer |
|
2014-06-18 |
| Architektura żywych budowli |
|
Yong |
|
2014-06-20 |
| Krótko żyjące zwierzęta i bardzo stare rośliny |
|
Zimmer |
|
2014-06-21 |
| Pająki społeczne wybierają swoje kariery |
|
Yong |
|
2014-06-23 |
| Skrzydlata rzeka |
|
Zimmer |
|
2014-06-25 |
| Dziwaczne, wysysające krew czerwie jurajskie |
|
Cobb |
|
2014-06-28 |
| Zaproszenie na wspólne polowanie |
|
Yong |
|
2014-06-30 |
| Marnie napisany artykuł o uroczym gryzoniu |
|
Coyne |
|
2014-07-03 |
| Jak przypadek pomógł znaleźć sposób na suszę |
|
Klein Leichman |
|
2014-07-04 |
| Przespać atak antybiotyku |
|
Yong |
|
2014-07-06 |
| Uprawy GM są dobre dla środowiska |
|
|
|
2014-07-08 |
| Trawa w uchu. Ale po co? |
|
Coyne |
|
2014-07-09 |
| Zoo w gębie |
|
Zimmer |
|
2014-07-10 |
| Suplementem diety wampira |
|
Yong |
|
2014-07-11 |
| Seks z wymarłym ludem dał gen życia na wysokości |
|
Yong |
|
2014-07-15 |
| Lot przez przestrzeń wewnętrzną |
|
Zimmer |
|
2014-07-17 |
| Osa, która zatyka wejście do gniazda trupami mrówek |
|
Yong |
|
2014-07-18 |
| Czym jest nauka i dlaczego ma nas obchodzić? |
|
Sokal |
|
2014-07-22 |
| Nowy, opierzony i czteroskrzydły dinosaur |
|
Coyne |
|
2014-07-23 |
| Oglądanie oceanu brzęczącym nosem |
|
Zimmer |
|
2014-07-26 |
| OLBRZYMI owad wodny (i kilka innych) |
|
Coyne |
|
2014-07-28 |
| Najbardziej zdumiewające oczy w przyrodzie |
|
Yong |
|
2014-07-29 |
| Czy jaszczurka “widzi” skórą |
|
Yong |
|
2014-08-02 |
| Nowy opierzony dinozaur sugeruje, że większość dinozaurów miała pióra |
|
Coyne |
|
2014-08-03 |
| Ewolucja łożyska a seksualna zimna wojna |
|
Yong |
|
2014-08-04 |
| Energia odnawialna nie działa |
|
Ridley |
|
2014-08-07 |
| Czy istnieje darwinowskie wyjaśnienie ludzkiej kreatywności? |
|
Dennett |
|
2014-08-08 |
| Gry zespołowe plemników |
|
Yong |
|
2014-08-09 |
| Oko ciemieniowe hatterii |
|
Mayer |
|
2014-08-10 |
| Osobisty mikrobiom w cyfrach |
|
Zimmer |
|
2014-08-14 |
| Izraelska koszulka EKG monitoruje serca, ratuje życie |
|
Shamah |
|
2014-08-17 |
| Grantowie na Galápagos i ich hybrydowe gatunki |
|
Coyne |
|
2014-08-18 |
| Ośmiornica dba o swoje jaja przez 53 miesiące, a potem umiera |
|
Yong |
|
2014-08-20 |
| Cuda genetyki: arbuz bez pestek |
|
Coyne |
|
2014-08-25 |
| Utracony sposób tworzenia ciał przed istnieniem szkieletów i muszli |
|
Yong |
|
2014-08-26 |
| Usunięcie obrzydzenia z medycyny mikrobiomowej |
|
Zimmer |
|
2014-08-28 |
| Tysiąc współpracujących, samorganizujących się robotów |
|
Yong |
|
2014-08-30 |
| Nogoprządki |
|
Naskręcki |
|
2014-09-01 |
| Drzewo zapachów |
|
Zimmer |
|
2014-09-02 |
| Sposób szczura na trujący pokarm |
|
Yong |
|
2014-09-05 |
| Raczkowanie w ewolucji |
|
Zimmer |
|
2014-09-06 |
| Zmieniająca kolor płachta zainspirowana skórą ośmiornicy |
|
Yong |
|
2014-09-08 |
| Erotyczna doniosłość bioder walenia |
|
Zimmer |
|
2014-09-11 |
| Co słychać w sprawie globalnego ocieplenia? |
|
Ridley |
|
2014-09-14 |
| Foki mogły przenieść gruźlicę do Nowego Świata |
|
Yong |
|
2014-09-16 |
| Jak kolibry odzyskały utracone przez ptaki odczuwanie słodyczy |
|
Yong |
|
2014-09-19 |
| Ochrona zagrożonych węży wymaga ochrony węży jadowitych |
|
Yong |
|
2014-09-22 |
| Uo, zaklinacz deszczu |
|
Naskręcki |
|
2014-09-23 |
| Co wypadające dyski mówią nam o 700 milionach lat ewolucji |
|
Zimmer |
|
2014-09-24 |
| O korzyściach przypadkowego kolekcjonowania okazów |
|
Naskręcki |
|
2014-09-28 |
| Pradawnym płazom odrastały kończyny |
|
Coyne |
|
2014-09-29 |
| Trawienny drapacz chmur |
|
Yong |
|
2014-09-30 |
| Ofiary naszych ułomności |
|
Naskręcki |
|
2014-10-02 |
| Jak dotarliśmy do teraźniejszości |
|
Ridley |
|
2014-10-05 |
| Dlaczego kod genetyczny nie jest uniwersalny |
|
Cobb |
|
2014-10-06 |
| Ukryte przed wzrokiem zoo w Central Park |
|
Zimmer |
|
2014-10-09 |
| Specjacja sympatryczna we wnętrzu cykady |
|
Yong |
|
2014-10-10 |
| Nocny stukot małych kopyt |
|
Naskręcki |
|
2014-10-12 |
| Wojna domowa w ludzkim genomie |
|
|
|
2014-10-13 |
| Penetrujący jaskinie robot-wąż wzorowany na grzechotnikach rogatych |
|
Yong |
|
2014-10-19 |
| Dowody ewolucji: wideo i nieco dłuższy wywód |
|
Coyne |
|
2014-10-22 |
| O wyższości lepszego nad gorszym |
|
Zimmer |
|
2014-10-26 |
| Powódź pożyczonych genów u powstania maleńkich ekstremistów |
|
Yong |
|
2014-10-30 |
| Tak, neandertalczycy to my! |
|
Mayer |
|
2014-11-04 |
| Zgarbowate |
|
Naskręcki |
|
2014-11-10 |
| Facet od nauki przeciwko GMO |
|
Coyne |
|
2014-11-12 |
| Figę dostaje ten kto rano wstaje |
|
Yong |
|
2014-11-13 |
| Mrówki, altruizm i poświęcenie |
|
Ridley |
|
2014-11-14 |
| Norowirus: doskonały patogen wyłania się z cienia |
|
Zimmer |
|
2014-11-15 |
| Siedem narzędzi myślenia |
|
Dennett |
|
2014-11-19 |
| Ciężarna wężyca przygotowuje się do macierzyństwa |
|
Yong |
|
2014-11-20 |
| Naturalność życia rodzinnego? |
|
Zimmer |
|
2014-11-25 |
| Genetyka kocich łat |
|
Coyne |
|
2014-11-26 |
| Świat RNA |
|
Cobb |
|
2014-11-27 |
| Dymorfizm płciowy i ideologia |
|
Coyne |
|
2014-12-01 |
| Gdy mutację przeciwstawić infekcji – od anemii sierpowatej do Eboli |
|
Lewis |
|
2014-12-02 |
| Nasze wewnętrzne pióra |
|
Zimmer |
|
2014-12-03 |
| Nie wszystkie muchy latają |
|
Naskręcki |
|
2014-12-04 |
| Jest tuż za tobą! Czy to duch, czy robot? |
|
Yong |
|
2014-12-06 |
| Najstarsza jak dotąd identyfikacja medycyny sądowej |
|
Coyne |
|
2014-12-10 |
| Samoloty bez pilotów i samochody bez kierowców |
|
Ridley |
|
2014-12-11 |
| Tworzenie dowodów w oparciu o politykę |
|
Ridley |
|
2014-12-16 |
| Pisklę przypominające wyglądem i zachowaniem trującą gąsienicę |
|
Coyne |
|
2014-12-18 |
| Wierzący nagradzani za życia |
|
Coyne |
|
2014-12-21 |
| Nietoperze owocożerne także mają sonar (ale niezbyt dobry) |
|
Yong |
|
2014-12-22 |
| Implanty WiFi do mózgu dla rąk robota |
|
Zimmer |
|
2014-12-25 |
| Naukowcy wprowadzają nową tradycję kulturową dzikim sikorkom |
|
Yong |
|
2014-12-26 |
| List do władz Uniwersytetu Harvarda |
|
Pinker |
|
2014-12-26 |
| Dlaczego zwierzęta są urocze? |
|
Coyne |
|
2014-12-30 |
| Dlaczego te dziwaczne owady sygnalizują ostrzeżenie po ataku? |
|
Yong |
|
2014-12-31 |
| Leniwce i pancerniki widzą czarno-biały świat |
|
Yong |
|
2015-01-06 |
| Ogony CAT osłabiają centralny dogmat – dlaczego ma to znaczenie i dlaczego nie ma |
|
Cobb |
|
2015-01-08 |
|
| « Poprzednia strona Następna strona » |
| | |