|
Moja ostatnia praca badawcza. Część 2: Wyniki
|
Jerry A. Coyne
| 2020-02-03
|
Kilka dni temu zacząłem dwuczęściowe podsumowanie (teraz stało się trzyczęściowe!) tego, co prawdopodobnie będzie moją ostatnią pracą „badawczą”: ostatnią pracą, w której własnymi rękami manipulowałem muszki Drosophila, by zebrać dane. (To nie znaczy, że będzie to mój ostatni artykuł naukowy.)
Ten post omawiał cele i metody naszego artykułu w „Genetics”, który możecie zobaczyć przez kliknięcie na link pod zrzutem z ekranu poniżej lub na pdf tutaj. Po napisaniu tego postu uznałem, że podzielę omówienie na trzy części, bo robi się to trochę zbyt długie. Dzisiaj mamy wyniki, a za kilka dni napiszę o tym, jakie jest moim zdaniem znaczenie tych wyników (tj. to, co nazywa się „dyskusją” w artykule”).
Przesłanki eksperymentu opisałem szczegółowo w poprzednim poście, więc nie będę się tu powtarzał. W skrócie, wzięliśmy dwie pary gatunków i dla każdej pary stworzyliśmy rój hybrydowy, składający się z osobników z połową genomu od każdego gatunku rodzicielskiego i połową organelli i cytoplazmy każdego gatunku rodzicielskiego (zawartości komórek). Każda para składała się z jednego szeroko rozpowszechnionego gatunku i jednego wyspiarskiego gatunku (D. simulans/D. mauritiana w jednym eksperymencie i D. yakuba/D. santomea w drugim). Celem było zobaczenie, czy rój hybrydowy powróci do jednego z gatunków rodzicielskich, pozostanie grupą „mieszańców”, które mają geny obu gatunków, lub może wyewoluuje w nowy laboratoryjny gatunek, który jest izolowany reprodukcyjnie od obu gatunków rodzicielskich.
Stworzyliśmy osiem replik „rojów” dla każdej pary gatunków, żeby zobaczyć, jak bardzo powtarzają się zmiany. I mierzyliśmy cztery zestawy cech, z których każdy charakteryzował i odróżniał dwa gatunki rodzicielskie:
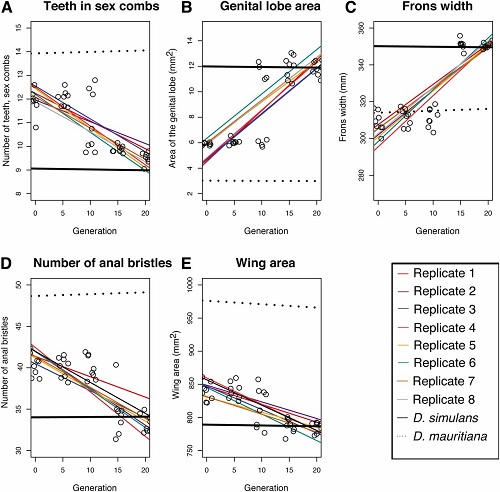
1.) Morfologiczne cechy, które odróżniały każdą parę gatunków. Było ich pięć w parze D. simulans/D. mauritiana i trzy w parze D. santomea/D. yakuba. Opisałem je w poprzednim poście, a pokazuje je także ilustracja poniżej. Cechy w roju (i w czystych gatunkach) mierzyliśmy co pięć pokoleń aż do pokolenia 20.
2.) Zachowanie godowe. Jak pisałem wcześniej, między członkami obu par jest dyskryminacja przy wyborze partnerów (i skrócona kopulacja). Wzięliśmy samce i samice z roju hybrydowego i kojarzyliśmy je z osobnikami z czystych gatunków rodzicielskich, żeby zobaczyć, czy hybrydy zachowywały się jak jeden lub drugi czysty gatunek (zrobiliśmy to w pokoleniu 21; utrzymywaliśmy roje przez 24 pokolenia).
Na przykład, kopulacje między samcami D. simulans i samicami D. mauritiana są bardzo krótkie w porównaniu do kopulacji wewnątrz czystych gatunków. Jeśli samce z roju hybrydowego także wykazują krótki czas trwania kopulacji z czystymi samicami D. mauritiana, możemy wnioskować, że ich cechy godowe powróciły do cech D. simulans. (Spodziewalibyśmy się także, że nie będą wykazywać dyskryminacji wobec D. simulans, ale znaczną niechęć do kopulacji z D. mauritiana.) Zrobiliśmy to dla obu par gatunków.
3.) Sterylność w krzyżówkach gatunków. Kiedy krzyżujesz członków obu par gatunków, otrzymujesz samce-hybrydy, które są sterylne i samice, które są płodne. (Ten wzór nazywa się „regułą Haldane’a). Sterylność trwa przez pewien czas w rojach hybrydowych, podczas gdy dobór usuwa geny dające sterylność. Mierzyliśmy sterylność samców w populacji w pokoleniu 20 w porównaniu do czystego gatunku (w którym samce są całkowicie płodne). Co ważniejsze, przyglądaliśmy się sterylności hybryd powstałych z krzyżówek między osobnikami z roju a dwoma czystymi gatunkami (wszystkie z pokolenia 20). Na przykład, jeśli skrzyżujesz rój D. mauritiana/D. simulans z gatunkami rodzicielskimi i otrzymasz płodne potomstwo z D. simulans, ale sterylne potomstwo z D. mauritiana, to wiesz, że genotypowy stan płodności w roju powrócił do stanu D. simulans.
4.) Skład DNA roju hybrydowego. Jak pisałem w poprzednim poście, DNA obu par gatunków różniły się w wielu miejscach. Mogliśmy więc sekwencjonować próbki roju hybrydowego i zobaczyć, jakie proporcje genomu każdego gatunku pozostały w roju po 20 pokoleniach. Zrobiliśmy to we wszystkich ośmiu replikach dla każdego z dwóch rojów, ale tylko raz, bo to przedsięwzięcie obejmowało sekwencjonowanie około 20 milionów zasad. Analiza jest trochę skomplikowana i nie będę się w nią zagłębiał tutaj.
Artykuł:
Poniżej podaję wyniki czterech testów. Wszystkie są zgodne i jednoznaczne: w każdej analizie, czy to morfologii, zachowania, czy sekwencji DNA rój powrócił do „dominującego” (kontynentalnego) gatunku: D. simulans w jednym roju i D. yakuba w drugim. I to samo było we wszystkich ośmiu replikach każdego roju. W pokoleniu 20 również sekwencje DNA wszystkich replik powróciły niemal całkowicie do dominującego gatunku, chociaż pozostało nieco DNA wyspiarskiego gatunku. Ten ”resztkowy” DNA był dość spójny we wszystkich replikach.
1.) Cechy morfologiczne. Dwa wykresy poniżej pokazują zmiany w morfologii cech odróżniających gatunki we wszystkich replikach rojów przez 20 pokoleń (mierzono pokolenia 0, 5, 10, 15 i 20). Pierwszy wykres pokazuje rój D. mauritiana/D. simulans a drugi rój D. yakuba/D. santomea. Mierzyliśmy także morfologię obu czystych gatunków jako grupy kontrolne w czasie; te wartości są prostymi i przerywanymi liniami na górze i na dole (były one w zasadzie stałe, bo są cechami specyficznymi dla gatunku). Hybrydowe repliki w każdym pokoleniu są pokazane jako otwarte koła, z średnimi między replikami wytyczonymi jako najlepiej pasująca linia, a każda replika ma inny kolor.
Na pierwszym wykresie poniżej wartości cech czystego gatunku D. mauritiana są liniami przerywanymi; wartości cech czystego D. simulans są liniami ciągłymi. Jak widać, rój hybrydowy w pokoleniu 0 jest w zasadzie pośredni między liniami, tak jak powinien być (szerokość czoła pozostaje jednak bliska czystemu D. mauritiana przez 10 pokoleń zanim powraca do wartości D. simulans ).
Ważnym wynikiem jest to, że z czasem każda replika pod względem każdej cechy powraca do wartości D. simulans. Zmiany są szczególnie zaznaczone między pokoleniami 10 a 15. W pokoleniu 20 wartości wszystkich cech są w zasadzie wartościami czystego D. simulans. Pod względem tych cech hybrydy wyewoluowały więc wszystkie wartości cech D. simulans. Rój wygląda jak czyste osobniki D. simulans.
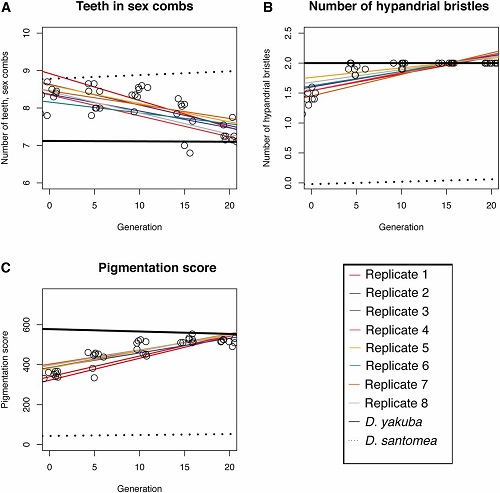
A to jest wykres dla cech roju D. yakuba/D. santomea, z wartościami czystego D. yakuba jako linia ciągła i wartościami D. santomea jako linia przerywana. Tutaj także cechy, które zaczęły się jako pośrednie w pierwszym pokoleniu stopniowo przyjmują wartości dominującego gatunku (D. yakuba) po 20 pokoleniach. Tak jest we wszystkich ośmiu replikach. W szczególności, pigmentacja muszek w roju staje się tak ciemna jak u czystych osobników D. yakuba (wynik około 600 na skali z maksimum 1200 punktów), zamiast niemal całkowitego braku pigmentacji jak D. santomea (ten czysty gatunek ma przeciętną liczbę punktów pigmentacji 49). Cechy zmieniają się w różnym tempie z upływem czasu. Na przykład, zmiana pigmentacji jest mniej więcej równa w czasie, ale zmiana w szczecinkach hypandrial powraca do wartości D. yakuba w ciągu 5 pokoleń. 2.) Zachowanie godowe. Mogę to streścić krótko:pod wszystkimi aspektami zalecania się i kopulacji rój hybrydowy powrócił do ”dominującego”, kontynentalnego gatunku we wszystkich replikach do pokolenia 21. Innymi słowy, w roju D. yakuba/D. santoma wszystkie osobniki zachowywały się jak czyste muszki D. yakuba, a w roju D. simulans/D. mauritiana wszystkie osobniki zachowywały się jak czyste D. simulans. To dotyczy to tak samo samców, jak samic. Mamy więc powrót do czystego gatunku nie tylko w morfologii, ale także w zachowaniu.
3.) Sterylność w krzyżówkach gatunków. Raz jeszcze we wszystkich replikach obu rojów hybrydowych osobniki powróciły do cech płodności ”dominującego”, kontynentalnego gatunku do pokolenia 20. Na przykład, patrząc na osobniki w 8. replice rojów D. simulans/D. mauritiana, wszystkie samce w roju miały płodnych męskich potomków kiedy krzyżowały się z samicami D. simulans, ale miały sterylnych męskich potomków, kiedy krzyżowały się z samicami D. mauritiana. Tak samo było, kiedy krzyżowaliśmy samice z rojów hybrydowych z samcami albo z D. simulans, albo z D. mauritiana. I tak samo było za samcami z roju D.santomea/D. yakuba: pod koniec eksperymentu zarówno samce z roju, jak samice z roju zachowywały się pod względem płodności tak samo jak osobniki czysto D. yakuba. Znowu obserwujemy powszechny i powtarzalny powrót do „dominującego”, kontynentalnego gatunku.
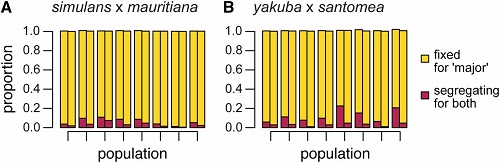
4.) Skład DNA roju hybrydowego. Test w pokoleniu 20 pokazał, że we wszystkich rojach w obu gatunkach DNA gatunku wyspiarskiego w zasadzie został wyeliminowany i genomy rojów były niemal całkowicie genomami dominujących gatunków. Jednak pewne resztki specyficzne dla gatunków wyspiarskich pozostały w obu replikach, jak tego oczekiwano, ponieważ część z tego jest „neutralna” i nie podlega selekcji. Tutaj, na przykład, jest proporcja przodków w każdej replice (dwa paski dla każdej repliki zależnie od tego, którego genomu używaliśmy jako odnośnika) z DNA dominującego gatunku na żółto i DNA wyspiarskiego gatunku na czerwono. Pozostało bardzo niewiele DNA specyficznego dla wyspiarskiego gatunku, a te resztki, które pozostają, są „segregujące” tj. miejsca, które mają jedną kopię DNA wyspiarskiego i jedną kopię DNA dominującego w każdej danej pozycji (proszę pamiętać, że są dwie kopie każdego genu). Wyraźnie, DNA gatunku wyspiarskiego jest na drodze do zniknięcia.
(From paper): Genetic ancestry rapidly and consistently regressed to that of one of the two parental species in all admixed populations. (A) The proportion of sites either fixing for D. simulans ancestry or still segregating for both parental species’ ancestry in each of the eight admixed D. mauritiana/simulans populations. (B) The proportion of sites either fixing for D. yakuba ancestry or still segregating for both parental species’ ancestry in each of the eight admixed D. santomea/yakuba populations. Sites were considered to still be segregating for both parental species’ ancestry if any of the ploidy = 8 genotypes 2 | 6 through 6 | 2 received a posterior probability >1/3. The left bar for each population summarizes results obtained when mapping to either the D. mauritiana (A) or the D. santomea reference genomes (B). Bars to the right, for each population, summarize results obtained when mapping to either the D. simulans (A) or D. yakuba (B) reference genomes.
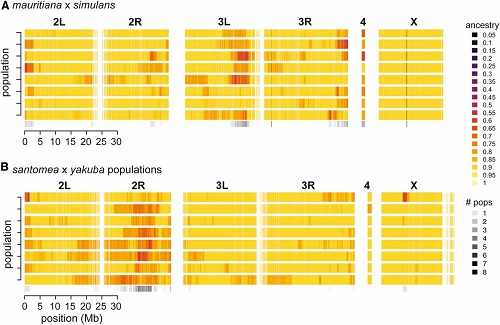
Tutaj jest szeroki skan genomu z każdą pionową linią reprezentującą okno na 5000 zasad ze składem gatunku zaznaczonym na żółto (DNA dominującego gatunku jest obecne) i czerwono (DNA wyspiarskiego gatunku jest obecne, ale z niezbyt wysoką częstotliwością). Czytając z góry do dołu, widać DNA na jednym ramieniu chromosomu w ośmiu replikach (istnieją cztery chromosomy z X mającym tylko po jednym ramieniu, drugim i trzecim po dwa ramiona i czwartym bardzo małym).
(From paper): Genome-wide distribution of ancestry in all admixed populations. Heatmaps showing ancestry estimates summarized in 5-kb genomic windows for each chromosome or chromosomal arm in the D. simulans (A) and D. yakuba (B) reference genomes. Each row is a different admixed population and colors reflect ancestry ranging from 0 (fixed for “minor” parent ancestry) to 1 (fixed for “major” parent ancestry). The bottom row summarizes the number of populations that showed evidence of a given genomic window still segregating for both parental species’ ancestry (i.e., ancestry estimate < 0.8).
Jak widać, olbrzymia większość genomu jest żółta, pochodząc z dominującego gatunku (czerwony kolor wybija się, ale nie występuje zbyt często na czysto żółtym tle).
Widać także, że pewne obszary genomu mają tendencję do pozostawania „segregującymi” we wszystkich replikach: na przykład, czubek trzeciego chromosomu w roju D. simulans/D. mauritiana i na środkowym ramieniu drugiego chromosomu w roju D. yakuba/ D. santomea.
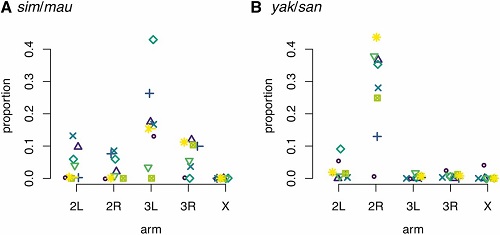
Wreszcie, żeby pokazać mizerną ilość obcego genomu, pozostającą w większości części genomu, tutaj jest wykres pokazujący dla każdego z pięciu ramion chromosomów, ile z genomu roju było segregującego: tj., jaka proporcja DNA miała pewne sekwencje od obu gatunków (niemal wszystko było „heterozygotyczne”, z jedną kopią sekwencji kontynentalnej i jedną kopią sekwencji wyspiarskiej). Osiem replik otrzymało różne symbole dla każdego ramienia.
Jak widać, dla niemal wszystkich miejsc chromosomów poza pewnymi replikami na ramieniu 3L w roju D. simulans/D. mauritiana i ramieniu 2R w roju D. yakuba/D. santomea, pozostało przeciętnie mniej niż 5% DNA gatunku wyspiarskiego. Na wszystkich ramionach poza jednym w roju D. yakuba/D. santomea nie pozostał niemal żaden obcy DNA.
Godne zauważenia jest to, że ilość obcego DNA na chromosomie X („płciowym”) była najniższa w obu rojach. To może być spowodowane faktem, że chromosom X zawiera szereg genów, które dają sterylność hybrydowych samców, a więc „wyspiarski” DNA został szybko wyeliminowany przez dobór. (A także, ponieważ X jest obecny tylko w jednej kopii u samców, zarówno recesywne, jak dominujące „geny sterylności” w pełni ulegają ekspresji, a więc są eliminowane dużo szybciej.)
(Caption from paper): The proportion of genomic windows where both parental species’ ancestry still segregated varied across chromosomes. Each point represents the proportion of 5-kb genomic windows that have evidence for both parental ancestries still segregating after 20 generations following initial hybridization between the parental species. (A) D. simulans/D. mauritiana; (B) D. yakuba/D. santomea.
Wielki wniosek: W replikach obu rojów i dla wszystkich mierzonych cech – morfologiczne, zachowania godowego, płodności i sekwencji DNA – cechy i DNA roju wyewoluowały (powróciły) w laboratorium do cech i DNA jednego, czystego gatunku. I we wszystkich wypadkach tym czystym gatunkiem był „dominujący”, kontynentalny gatunek: D. simulans w jednym roju i D. yakuba w drugim. Nie otrzymaliśmy „hybrydowego” gatunku, ale raczej dostaliśmy populację, której DNA był w zasadzie DNA czystego gatunku. Wyniki były wyraźnie zgodne we wszystkich replikach i ogólne wyniki bardzo podobne w obu rojach.
W następnym (i ostatnim) poście spróbuję opisać znaczenie tych wyników i przedstawić pewne teorie o tym, dlaczego we wszystkich wypadkach nasze roje powróciły do dominującego, nie zaś wyspiarskiego gatunku.
Jeśli dotarliście tak daleko, dziękuję za przeczytanie!
________________ Matute, D. R., A. A. Comeault, E. Earley, A. Serrato-Capuchina, D. Peede, A. Monroy-Eklund, W. Huang, C. D. Jones, T. F. C. Mackay, and J. A. Coyne. 2020. Rapid and predictable evolution of admixed populations between two Drosophila species pairs. Genetics 214:211-230.
My last research paper. Part 2 Results
Why Evolution Is True, 28 stycznia 2020 Tłumaczenie: Małgorzata Koraszewska Jerry A. CoyneEmerytowany profesor na wydziale ekologii i ewolucji University of Chicago, jego książka "Why Evolution is True" (Polskie wydanie: "Ewolucja jest faktem", Prószyński i Ska, 2009r.) została przełożona na kilkanaście języków, a przez Richarda Dawkinsa jest oceniana jako najlepsza książka o ewolucji. Jerry Coyne jest jednym z najlepszych na świecie specjalistów od specjacji, rozdzielania się gatunków. Jest również jednym ze znanych "nowych ateistów" i autorem książki "Faith vs Fakt". Jest wielkim miłośnikiem kotów i osobistym przyjacielem redaktor naczelnej.
Znalezionych 1479 artykuły.
|
|
|
|
|
|
|
| Opadający liść, latający smok |
|
Yong |
|
2015-01-10 |
| Nowotwory są konsekwencją wieku, a nie grzechu |
|
Ridley |
|
2015-01-11 |
| Lekcja ewolucji: specjacja w akcji! |
|
Coyne |
|
2015-01-12 |
| Epidemiologia |
|
Feldman |
|
2015-01-13 |
| Aquilops, mały dinozaur, który wiele mógł |
|
Farke |
|
2015-01-15 |
| Mózgi dwudysznych wcale nie są nudne |
|
Farke |
|
2015-01-18 |
| Nasi przyjaźni rozkładacze drożdży |
|
Yong |
|
2015-01-19 |
| Rok 2014 był świetny dla Hupehsuchia |
|
Farke |
|
2015-01-24 |
| Czy mikrobiom może się zbuntować? |
|
Zimmer |
|
2015-01-28 |
| Moje życie zwolennika łagodnego ocieplenia |
|
Ridley |
|
2015-01-29 |
| Dan Brown - akomodacjonista |
|
Coyne |
|
2015-01-31 |
| Towarzyskim małpom w zimie jest cieplej |
|
Yong |
|
2015-02-01 |
| Miejsce dla Hallucigenii |
|
Łopatniuk |
|
2015-02-08 |
| Frankenstein dziś nie może wyjść i się bawić |
|
Zimmer |
|
2015-02-11 |
| Skaczący DNA i ewolucja ciąży |
|
Yong |
|
2015-02-12 |
| Mitochondrialna donacja jest cudowną możliwością |
|
Ridley |
|
2015-02-13 |
| O pochodzeniu kolorowych twarzy małp |
|
Yong |
|
2015-02-16 |
| Mimikra chemiczna u mszyc |
|
Coyne |
|
2015-02-19 |
| Ogon ćmy i nietoperze |
|
Coyne |
|
2015-02-23 |
| Nasze wewnętrzne wirusy: obecne od 40 milionów lat |
|
Zimmer |
|
2015-02-27 |
| Jak wirus odry stał się mistrzem zarażania |
|
Zimmer |
|
2015-03-01 |
| Łowienie mikrobów u podstaw niedożywienia |
|
Yong |
|
2015-03-03 |
| Astrocyty tworzą nowe neurony po udarze |
|
Łopatniuk |
|
2015-03-04 |
| Trzecia droga ewolucji? Nie sądzę |
|
Coyne |
|
2015-03-05 |
| Nie igraj z odrą |
|
Łopatniuk |
|
2015-03-06 |
| Myszy z wszczepionym ludzkim DNA mają większe mózgi |
|
Yong |
|
2015-03-09 |
| Pasożytnicze osy zarażone kontrolującymi umysł wirusami |
|
Zimmer |
|
2015-03-10 |
| Twój spadek po przodkach, drogi strunowcu |
|
Łopatniuk |
|
2015-03-12 |
| Modliszka storczykowa: czy upodabnia się do storczyka? |
|
Coyne |
|
2015-03-13 |
| Ebola przenoszona drogą kropelkową? |
|
Zimmer |
|
2015-03-17 |
| Woda odskakuje od skóry gekona |
|
Yong |
|
2015-03-19 |
| Czerwonogłowe muchy |
|
Naskręcki |
|
2015-03-22 |
| Porywacze mitochondriów |
|
Łopatniuk |
|
2015-03-23 |
| Jesteśmy błyskawicznymi rozgryzaczami liczb |
|
Zimmer |
|
2015-03-24 |
| Seks paproci i kreacjoniści |
|
Coyne |
|
2015-03-27 |
| Piersi i jajniki, czyli rak i święto błaznów |
|
Łopatniuk |
|
2015-03-28 |
| Walenie po niewłaściwej stronie świata |
|
Zimmer |
|
2015-03-31 |
| Paliwa kopalne nie są wyczerpane, nie są przestarzałe, nie są złe |
|
Ridley |
|
2015-04-01 |
| Francis Crick był niesamowitym geniuszem |
|
Coyne |
|
2015-04-02 |
| Matrioszki, czyli płód w płodzie (fetus in fetu) |
|
Łopatniuk |
|
2015-04-03 |
| Jak ryba łyka pokarm na lądzie? |
|
Yong |
|
2015-04-04 |
| Dobór krewniaczy pozostaje wartościowym narzędziem |
|
Coyne |
|
2015-04-06 |
| Malaria pachnąca cytryną |
|
Zimmer |
|
2015-04-07 |
| Nowotwory sprzed tysiącleci |
|
Łopatniuk |
|
2015-04-08 |
| Nowa i dziwaczna, zmieniająca kształt żaba |
|
Coyne |
|
2015-04-10 |
| Czy mleko matek może odżywiać manipulujące umysłem mikroby? |
|
Yong |
|
2015-04-14 |
| Wczesna aborcja farmakologiczna – skuteczna i bezpieczna, a w Arizonie w dodatku – odwracalna |
|
Łopatniuk |
|
2015-04-15 |
| Małpo ty moja |
|
Koraszewski |
|
2015-04-17 |
| Jak często geny przeskakują między gatunkami? |
|
Coyne |
|
2015-04-18 |
| Młode mysie matki i oksytocyna |
|
Yong |
|
2015-04-21 |
| Ciąg dalszy sporu o dobór grupowy |
|
Coyne |
|
2015-04-22 |
| Jak psy zdobywają nasze serca? |
|
Yong |
|
2015-04-23 |
| Niebo gwiaździste nade mną |
|
Łopatniuk |
|
2015-04-24 |
| Żywotne pytanie |
|
Ridley |
|
2015-04-25 |
| Czy rozum jest “większy niż nauka”? Kiepska próba deprecjonowania nauki |
|
Coyne |
|
2015-04-28 |
| Kiedy Darwin spotkał inną małpę |
|
Zimmer |
|
2015-04-30 |
| Redagowanie ludzkich embrionów: Pierwsze próby |
|
Zimmer |
|
2015-05-04 |
| Robaki i rak |
|
Łopatniuk |
|
2015-05-09 |
| Nowe skamieniałości: najwcześniejszy na świecie znany ptak |
|
Coyne |
|
2015-05-12 |
| Pradawny DNA czyni z prehistorii otwartą książkę |
|
Ridley |
|
2015-05-13 |
| Chiński dinozaur miał skrzydła jak nietoperz i pióra |
|
Yong |
|
2015-05-14 |
| Czy człowiek musiał wyewoluować? |
|
Coyne |
|
2015-05-15 |
| Gigantyczne walenie mają super elastyczne nerwy |
|
Yong |
|
2015-05-18 |
| Znikające badaczki, czyli Sophie Spitz była kobietą |
|
Łopatniuk |
|
2015-05-21 |
| Bambusowi matematycy |
|
Zimmer |
|
2015-05-25 |
| Pierwsza znana ryba ciepłokrwista |
|
Coyne |
|
2015-05-27 |
| Puszek kłębuszek, zdobywca serduszek |
|
Łopatniuk |
|
2015-05-28 |
| Jak powiększyć kapitał naturalny |
|
Ridley |
|
2015-05-30 |
| Symbiotyczna katastrofa długoletniej cykady |
|
Yong |
|
2015-06-02 |
| Przypuszczalnie złamana kość |
|
Coyne |
|
2015-06-04 |
| Tajemnica kangurzych adopcji |
|
Zimmer |
|
2015-06-05 |
| Proszalne mruczenie kota zawiera płacz, dźwięk bardziej naglący i nieprzyjemny niż normalne mruczenie |
|
Coyne |
|
2015-06-09 |
| Jak afrykańskie obszary trawiaste utrzymują tak wiele roślinożernych? |
|
Yong |
|
2015-06-11 |
| Co tam, panie, w anatomii, czyli mózg, naczynia limfatyczne i inne drobiazgi |
|
Łopatniuk |
|
2015-06-13 |
| Uratujmy producentów zombi! |
|
Zimmer |
|
2015-06-15 |
| Mikrob, który dokonał inwazji karaibskich raf koralowych |
|
Yong |
|
2015-06-16 |
| Ekomodernizm i zrównoważona intensyfikacja |
|
|
|
2015-06-17 |
| Kości! Wszędzie kości! |
|
Łopatniuk |
|
2015-06-20 |
| Cud? Ryba-piła urodzona z dziewiczej matki |
|
Coyne |
|
2015-06-23 |
| Rozproszony potencjał umysłowy owadów społecznych |
|
Yong |
|
2015-06-27 |
| Jak i dlaczego ta gąsienica gwiżdże? |
|
Coyne |
|
2015-06-30 |
| Co mamy zrobić z neuroróżnorodnością? |
|
Coyne |
|
2015-07-02 |
| Ser z czekoladą, czyli w kuchni u patologów |
|
Łopatniuk |
|
2015-07-04 |
| Nadajniki GPS zapowiadają nową epokę w badaniu zachowań zwierząt |
|
Yong |
|
2015-07-06 |
| Seksizm w nauce: czy Watson i Crick naprawdę ukradli dane Rosalind Franklin? |
|
Cobb |
|
2015-07-07 |
| Pielęgnice z jeziora w Kamerunie prawdopodobnie nie podlegały specjacji sympatrycznej: Część 1 |
|
Coyne |
|
2015-07-09 |
| Pielęgnice z jeziora w Kamerunie prawdopodobnie nie podlegały specjacji sympatrycznej: Część 2 |
|
Coyne |
|
2015-07-10 |
| Nowotwory spoza pakietu, czyli nie tylko czerniak |
|
Łopatniuk |
|
2015-07-11 |
| Photoshop czy nie photoshop? |
|
Naskręcki |
|
2015-07-13 |
| Gatunki inwazyjne są największym powodem wymierania |
|
Ridley |
|
2015-07-14 |
| Depresja inbredowa u człowieka |
|
Mayer |
|
2015-07-15 |
| Rozmowy między dzbanecznikiem a nietoperzem |
|
Yong |
|
2015-07-16 |
| Zdumiewająca historia dwóch par bliźniąt |
|
Coyne |
|
2015-07-17 |
| Ten chrząszcz niszczy twoją kawę przy pomocy bakterii |
|
Yong |
|
2015-07-22 |
| Co wojny o klimat zrobiły nauce |
|
Ridley |
|
2015-07-23 |
| Zabójcy z bagien |
|
Naskręcki |
|
2015-07-25 |
| Jak olbrzymie krewetki mogą zwalczać chorobę tropikalną i biedę |
|
Yong |
|
2015-07-28 |
| Ostrogony nie są naprawdę “żywymi skamieniałościami” |
|
Coyne |
|
2015-07-29 |
| Czworonożny wąż |
|
Mayer |
|
2015-07-30 |
| Gwałtownie ocieplający się klimat wywołał rewolucję megafauny |
|
Yong |
|
2015-07-31 |
|
| « Poprzednia strona Następna strona » |
| | |