Jak wyewoluowało ubarwienie ostrzegawcze?
|
Jerry A. Coyne
| 2023-06-12
|
Moja żaba, Atelopus coynei, także rzuca się w oczy i może być toksyczna, ale niezbyt wiele o tym wiemy
Ubarwienie aposematyczne, często nazywane „ubarwieniem ostrzegawczym”, to obecność jasnych lub rzucających się w oczy kolorów lub wzorów u zwierząt, które są toksyczne, szkodliwe, niebezpieczne lub trujące dla drapieżników. Oto przykład z
Wikipedii,
drzewołaz ziarnisty (
Oophaga granulifera). Podobnie jak wiele żab drzewołazowatych,
ta ma w swojej skórze wiele trujących alkaloidów i były one używane w Ameryce Środkowej i Południowej do pokrywania ostrzy strzał lub rzutek, które mogą zabijać ssaki. Każdy drapieżnik, który próbowałby zjeść jedną z takich żab, prawdopodobnie byłby martwy lub przynajmniej bardzo chory.
Atelopus coynei. Zdjęcie: Jordy Salazar/EcoMinga
Ale oczywiście nie tylko płazy są aposematyczne. Skunks reklamuje swoją toksyczność parą wyraźnych pasków. Wiele owadów, takich jak biedronki i niektóre motyle, jest również aposematycznych i toksycznych, w tym co najmniej jeden gatunek ptaków: zobacz tutaj, aby znaleźć obrazy zwierząt aposematycznych w Google.
Kolory i wzory dają ich nosicielom przewagę ewolucyjną nad ich przypuszczalnie zakamuflowanymi przodkami, ponieważ drapieżniki celowo unikają wzoru, zwykle dlatego, że nauczyły się go rozpoznawać i trzymać się od niego z daleka z powodu wcześniejszych nieprzyjemnych doświadczeń. (Unikanie może również wyewoluować, zamiast wyuczenia. Nawet jeśli zjedzenie jednego z nich zabije, preferowane będą te indywidualne drapieżniki, które mają mniejszą skłonność do atakowania wzorca). Zazwyczaj jednak jest to wyuczone.
Jednak, by uzyskać tę przewagę, gatunek aposematyczny musi być wystarczająco liczny, aby dać drapieżnikom szansę na nauczenie się, a następnie unikanie kolejnego zwierzęcia aposematycznego. A to stwarza problem ewolucyjny.
Jesteśmy prawie pewni, że gatunki aposematyczne wyewoluowały z gatunków zakamuflowanych. Aby rozpoczęło się ostrzegawcze zabarwienie, w zakamuflowanej populacji muszą wystąpić mutacje, które wytworzą osobniki o jasnych kolorach i wzorach, przynajmniej w początkowej formie.
I na tym polega problem: pierwszy zmutowany osobnik jest bardziej narażony na zaatakowanie i zabicie niż zakamuflowane osobniki. Nawet jeśli jest toksyczny, może zostać zabity lub zraniony, ponieważ jest nowym, rzucającym się w oczy stworzeniem. Jak więc adaptacja rozprzestrzenia się w populacji z rzadkiego stanu początkowego?
Wcześniej, jak opisano w doskonałym podsumowaniu Nature News & Views autorstwa Tima Caro poniżej (kliknij, aby przeczytać), mieliśmy kilka odpowiedzi:
1.) Cecha mogła wyewoluować w wyniku doboru krewniaczego u zwierząt stadnych. Chociaż pierwszy zmutowany osobnik może zostać zaatakowany, może należeć do grupy krewnych, którzy podzielają tę mutację aposematyczną. Zakładając, że drapieżnik nauczy się unikać wzorca po zabiciu lub zranieniu pierwszego osobnika, będzie unikać jego krewnych o podobnym kolorze, a to jest forma doboru krewniaczego genów koloru/wzoru, która może sprawić, że się rozprzestrzenią.
2.) Cecha mogła wyewoluować ze stanu, który był rzucający się w oczy, ale nie tak rzucający się w oczy, jak zwierzęta powyżej. Ale to prowadzi do tego samego problemu, co punkcie 1!
3.) Zaatakowany mutant aposematyczny mógłby uniknąć zabicia przez drapieżnika, ponieważ źle pachnie lub smakuje lub jest tylko lekko ranny. Jeśli drapieżnik uczy się na podstawie jednego doświadczenia (a niektóre tak robią), wówczas ten osobnik byłby odtąd chroniony przed drapieżnictwem, być może dając przewagę zmutowanemu genowi koloru/wzoru. Wydaje się to dość prawdopodobne i można to sprawdzić, podsuwając niedoświadczonym drapieżnikom aposematyczną zdobycz.
4.) Drapieżniki mogą generalnie unikać nowych kolorów lub wzorów, ponieważ mają obraz wyszukiwania gatunków jadalnych. Jak mówi Caro, są na to również pewne dowody.
Obecnie, w swoim podsumowaniu oryginalnej pracy, Caro opisuje piątą hipotezę, która jest opisana w artykule „Science” poniżej. Autorzy testują tę interesującą hipotezę za pomocą danych filogenetycznych i wydaje się, że jest ona potwierdzona.
Kliknij na link do oryginalnego artykułu w „Science” poniżej, aby przeczytać o nowej hipotezie ewolucji aposematyzmu. Autorzy testują to na płazach, ale może to dotyczyć również innych stworzeń. Możesz również znaleźć pdf tutaj.
Hipoteza autorów, która jest bardzo sprytna, jest taka, że pełne aposematyczne ubarwienie mogło wyewoluować, przynajmniej u płazów z wcześniejszego stanu, w którym nie było wyraźnie widoczne dla drapieżników. Może to obejmować kolory/wzory rozpoczynające swoją ewolucję na DOLNEJ (brzusznej) stronie zwierzęcia, która nie zwróciłaby uwagi, dopóki zwierzę nie zostało zaatakowane, w którym to momencie mogłoby błysnąć swoim wzorem i prawdopodobnie przestraszyć drapieżnika (drapieżnik może również dowiedzieć się z krótkiego spotkania, że ofiara była toksyczna). A sam kolor brzusznej strony może być dwojakiego rodzaju: małe plamy na powierzchni brzusznej (PV) lub w pełni zabarwiona powierzchnia brzuszna (FV). Kontrastuje to ze zwierzęciem, które jest w pełni ubarwione na całym ciele.
Gdy drapieżnik zaczął uczyć się, co oznacza kolor/wzór od zwierząt, które miały go na stronie brzusznej, kolor mógł ewoluować, aby pokryć zwierzę, czyniąc je w pełni aposematycznym.
Ale jak sprawdzić tę hipotezę? Cóż, można zobaczyć, czy drapieżniki nauczą się unikać toksycznych płazów, które miały kolorowe plamy namalowane na brzuchu, ale niewiele jest płazów, które są toksyczne i nie mają aposematycznego ubarwienia. Nie, autorzy sprawdzili swoją hipotezę, przeprowadzając rekonstrukcję filogenetyczną: wykorzystali żywe gatunki i ich znane drzewo genealogiczne, aby wywnioskować, jaki był kolor/wzór przodków. Ten rodzaj rekonstrukcji, który ma sens, jeśli masz wystarczającą ilość danych, jest coraz częściej wykorzystywany do badania ewolucji.
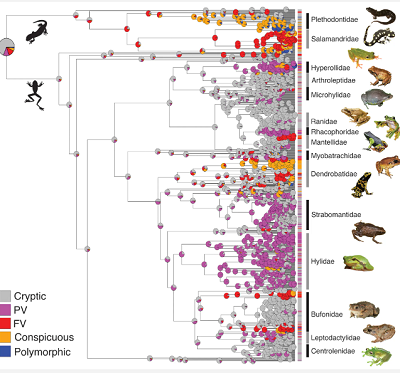
I tak Loeffler-Henry i in. dokonali wielkiej rekonstrukcji ewolucyjnej historii płazów, z których wiele było aposematycznie ubarwionych. Użyli 1106 gatunków, umieszczając każdy z nich w jednej z pięciu kategorii ewolucyjnych:
gatunek zakamuflowany („cry” na zdjęciu poniżej)
gatunek PV (strona brzuszna częściowo aposematyczna)
gatunek FV (strona brzuszna całkowicie aposematyczna)
gatunek całkowicie aposematyczny na całym ciele („conspicuous” lub „con” na zdjęciu poniżej)
gatunek polimorficzny (niektóre osobniki są aposematyczne, inne nie). Nie ma ich wiele i nie będę wchodził w to, dlaczego zakłada się ich istnienie.
Oto zdjęcie z artykułu przedstawiające cztery z pięciu stanów (nie pokazano gatunku polimorficznego):
Część podpisu z artykułu: Cry: zakamuflowany; PV (częściowo widoczna strona brzuszna): zakamuflowany grzbiet z wyraźnym kolorem występującym w postaci małych plam na normalnie ukrytych częściach ciała; FV (w pełni rzucająca się w oczy strona brzuszna): zakamuflowany grzbiet z wyraźnymi kolorami całkowicie pokrywającymi stronę brzuszną; Con: rzucający się w oczy.
A oto rekonstrukcja filogenezy pokazująca pozycję na drzewie genealogicznym każdego z pięciu stanów.
(Z artykułu): Ryc. 2. Oszacowanie stanu każdego stanu ubarwienia przodków (N = 1106 gatunków) u żab i salamander. Wykresy kołowe w każdym węźle pokazują prawdopodobieństwa stanów przodków. Stan przodków żab i salamander prawdopodobnie ma kamuflujące ubarwienie. Ukryte sygnały koloru (PV i FV) są szeroko rozpowszechnione i ewoluowały wielokrotnie w różnych liniach. PV: zakamuflowany grzbiet z rzucającym się w oczy kolorem występującym jako małe plamki na normalnie ukrytych częściach ciała; FV: zakamuflowany grzbiet z wyraźnymi kolorami całkowicie pokrywającymi stronę brzuszną. Zobacz tabelę S11, aby uzyskać informacje o autorach zdjęć.
W każdym węźle drzewa znajduje się diagram kołowy pokazujący prawdopodobieństwo, że ten przodek miał jeden z pięciu ocenionych stanów. Nie będę wchodził w metody wyprowadzania prawdopodobieństw (prawda, nie rozumiem ich); ale tutaj są najistotniejsze punkty:
1.) Przodkowie są zwykle mało wyróżniający się (zakamuflowani; szare kropki), z możliwym wyjątkiem niektórych salamander. Jest to zgodne z ewolucyjnym poglądem, że aposematyczne ubarwienie nie było stanem przodków, ale wyewoluowało jako adaptacja obronna mająca na celu odstraszanie drapieżników.
2.) Pełny aposematyzm – stan pomarańczowy – pojawił się dopiero później u płazów i
3.) . . . na ogół działo się to przez przejście przez pośredni stan aposematycznego ubarwienia na brzuchu (gatunki fioletowe i czerwone)
4.) Przewaga fioletowych kółek wcześniej niż czerwonych sugeruje, że stan pełnego ubarwienia brzusznego był poprzedzony w czasie ewolucją częściowego zabarwienia brzusznego: kolorowe plamy, które mogły być błyskawicznie pokazywane, ale wciąż są mniej widoczne dla drapieżników niż pełne zabarwienie strony brzusznej. Ta sugestia jest poparta analizą statystyczną prawdopodobieństwa modeli, ale pominę to.
To jest analiza płazów, ale równie dobrze może odnosić się do innych gatunków. W rzeczywistości wiele motyli, które mają ostrzegawcze ubarwienie, ma je na tylnych skrzydłach, które są zakryte podczas odpoczynku. Dopiero gdy latają lub przestraszy je drapieżnik, ujawnia się aposematyczne ubarwienie. Oto przykład: aposematyczny motyl z Nature Notes Raya Cannona. To motyl ptasznik pospolity (Troides helena), o którym wiadomo, że jest bardzo trujące, ponieważ larwy żywią się roślinami zawierającymi toksyczne kwasy arystolochowe.
(ze strony): Altinote dicaeus callianira – jego wyraźny wzór informuje o jego niejadalności. Zdjęcie: Adrian Hoskins
Przez długi czas ewolucja ubarwienia aposematycznego przedstawiała problem nazywany przez ewolucjonistów „doliną adaptacyjną”: jak przejść od jednego stanu adaptacyjnego (kamuflażu) do stanu przypuszczalnie lepiej przystosowanego (aposematyzmu), kiedy pośrednie stadium ewolucyjne (pierwszy zmutowany osobnik) był w niekorzystnej sytuacji: pogrążony w adaptacyjnej dolinie? Nie mogło to nastąpić w wyniku doboru naturalnego, ponieważ dobór nie może faworyzować osobników mniej przystosowanych (tutaj „rzadziej unikanych”).
Autorzy proponują rozwiązanie tego problemu: dolina adaptacyjna nie została przekroczona, ponieważ stan pośredni - ubarwienie brzuszne - dawał selektywną przewagę pierwszym zmutowanym osobnikom.
Autorzy kończą artykuł sugestią, że ich scenariusz mógłby dotyczyć wielu gatunków:
. . . makroewolucyjne badania nad ubarwieniem zwierząt powinny uwzględniać te niedoceniane ukryte sygnały, które są zarówno powszechne, jak i szeroko rozpowszechnione w królestwie zwierząt, aby pogłębić naszą wiedzę na temat ewolucji mechanizmów obronnych drapieżników. Rzeczywiście, wiele taksonów zwierząt, takich jak węże, ryby i różne stawonogi (patrz przykładowe grupy na ryc. S12) obejmuje gatunki, które są zakamuflowane, są aposematyczne i mają ukryte wyraźne sygnały. Dlatego zachęcamy do dalszych badań w innych taksonach, aby ocenić ogólność hipotezy szczebli jako drogi do aposematyzmu.
_________________
Loeffler, K., C. Kang, and T. N. Sherratt. 2023. Evolutionary transitions from camouflage to aposematism: Hidden signals play a pivotal role. Science 379:1136-1140. DOI: 10.1126/science.ade5156
Link do oryginału: https://whyevolutionistrue.com/2023/06/05/how-did-warning-coloration-evolve/
Why Evolution Is True, 5 czerwca 2023
Tłumaczenie: Małgorzata Koraszewska
Jerry A. Coyne
Emerytowany profesor na wydziale ekologii i ewolucji University of Chicago, jego książka "Why Evolution is True" (Polskie wydanie: "Ewolucja jest faktem", Prószyński i Ska, 2009r.) została przełożona na kilkanaście języków, a przez Richarda Dawkinsa jest oceniana jako najlepsza książka o ewolucji. Jerry Coyne jest jednym z najlepszych na świecie specjalistów od specjacji, rozdzielania się gatunków. Jest również jednym ze znanych "nowych ateistów" i autorem (wydanej również po polsku przez wydawnictwo "Stapis") książki "Faith vs Fakt". Jest wielkim miłośnikiem kotów i osobistym przyjacielem redaktor naczelnej.
Komentarze
Znalezionych 1477 artykuły.
|
|
|
|
|
|
|
| Kameleon przekazuje różne informacje różnymi częściami ciała |
|
Yong |
|
2013-12-14 |
| Paradoksalne cechy genetyki inteligencji |
|
Ridley |
|
2013-12-18 |
| Wielki skandal z biopaliwami |
|
Lomborg |
|
2013-12-19 |
| Przedwczesna wiadomość o śmierci samolubnego genu |
|
Coyne |
|
2013-12-22 |
| Czy jest życie na Europie? |
|
Ridley |
|
2013-12-22 |
| Nowa data udomowienia kotów: około 5300 lat temu – i to w Chinach |
|
Coyne |
|
2013-12-26 |
| Na Zeusa, natura jest przeżarta rują i korupcją |
|
Koraszewski |
|
2013-12-26 |
| Proces cywilizacji |
|
Ridley |
|
2013-12-28 |
| Jak karakara wygrywa z osami |
|
Cobb |
|
2013-12-29 |
| Żebropławy, czyli dziwactwa ewolucji |
|
Coyne |
|
2013-12-30 |
| Czy może istnieć sztuka bez artysty? |
|
Wadhawan |
|
2013-12-30 |
| Zderzenie mentalności |
|
Koraszewski |
|
2014-01-01 |
| Skrzydlaci oszuści i straż obywatelska |
|
Young |
|
2014-01-02 |
| Delfiny umyślnie narkotyzują się truciznami rozdymków |
|
Coyne |
|
2014-01-04 |
| Długi cień anglosfery |
|
Ridley |
|
2014-01-05 |
| Ciemna materia genetyki psychiatrycznej |
|
Zimmer |
|
2014-01-06 |
| Co czyni nas ludźmi? |
|
Dawkins |
|
2014-01-07 |
| Twoja choroba na szalce |
|
Yong |
|
2014-01-08 |
| Czy mamut włochaty potrzebuje adwokata? |
|
Zimmer |
|
2014-01-09 |
| Pradawne rośliny kwitnące znalezione w bursztynie |
|
Coyne |
|
2014-01-10 |
| Ratując gatunek możesz go niechcący skazać |
|
Yong |
|
2014-01-11 |
| Ewolucja ukryta w pełnym świetle |
|
Zimmer |
|
2014-01-13 |
| Koniec humanistyki? |
|
Coyne |
|
2014-01-15 |
| Jak poruszasz nogą, która kiedyś była płetwą? |
|
Yong |
|
2014-01-16 |
| Jak wyszliśmy na ląd, kość za kością |
|
Zimmer |
|
2014-01-19 |
| Twoja wewnętrzna mucha |
|
Cobb |
|
2014-01-22 |
| Ukwiał żyje w antarktycznym lodzie! |
|
Coyne |
|
2014-01-25 |
| Dlaczego poligamia zanika? |
|
Ridley |
|
2014-01-26 |
| Wspólne pochodzenie sygnałów płodności |
|
Cobb |
|
2014-01-28 |
| Ewolucja i Bóg |
|
Coyne |
|
2014-01-29 |
| O delfinach, dużych mózgach i skokach logiki |
|
Yong |
|
2014-01-30 |
| Dziennikarski „statek upiorów” Greg Mayer |
|
Mayer |
|
2014-01-31 |
| Dlaczego leniwce wypróżniają się na ziemi? |
|
Bruce Lyon |
|
2014-02-02 |
| Moda na kopanie nauki |
|
Coyne |
|
2014-02-03 |
| Neandertalczycy: bliscy obcy |
|
Zimmer |
|
2014-02-05 |
| O pochodzeniu dobra i zła |
|
Coyne |
|
2014-02-05 |
| Sposób znajdowania genów choroby |
|
Yong |
|
2014-02-07 |
| Czy humaniści boją się nauki? |
|
Coyne |
|
2014-02-07 |
| Kiedy zróżnicowały się współczesne ssaki łożyskowe? |
|
Mayer |
|
2014-02-10 |
| O przyjaznej samolubności |
|
Koraszewski |
|
2014-02-12 |
| Skąd wiesz, że znalazłeś je wszystkie? |
|
Zimmer |
|
2014-02-15 |
| Nauka odkrywa nową niewiedzę o przeszłości |
|
Ridley |
|
2014-02-18 |
| Żyjące gniazdo? |
|
Zimmer |
|
2014-02-19 |
| Planeta tykwy pospolitej |
|
Zimmer |
|
2014-02-21 |
| Nowe niezwykłe skamieniałości typu “Łupki z Burgess” |
|
Coyne |
|
2014-02-22 |
| Dziennik z Mozambiku: Pardalota |
|
Naskręcki |
|
2014-02-23 |
| Wskrzeszona odpowiedź z kredy na “chorobę królów” |
|
Yong |
|
2014-02-26 |
| Dziennik z Mozambiku: Sybilla |
|
|
|
2014-03-01 |
| Spojrzeć ślepym okiem |
|
Yong |
|
2014-03-02 |
| Intelektualne danie dnia The Big Think |
|
Coyne |
|
2014-03-04 |
| Przeczołgać się przez mózg i nie zgubić się |
|
Zimmer |
|
2014-03-05 |
| Gdzie podziewają się żółwiki podczas zgubionych lat? |
|
Yong |
|
2014-03-10 |
| Supergen, który maluje kłamcę |
|
Yong |
|
2014-03-14 |
| Idea, którą pora oddać na złom |
|
Koraszewski |
|
2014-03-15 |
| Zwycięstwa bez chwały |
|
Ridley |
|
2014-03-17 |
| Twarde jak skała |
|
Naskręcki |
|
2014-03-18 |
| Pasożyty informacyjne |
|
Zimmer |
|
2014-03-19 |
| Seymour Benzer: humor, historia i genetyka |
|
Cobb |
|
2014-03-21 |
| Kto to był Per Brinck? |
|
Naskręcki |
|
2014-03-23 |
| Potrafimy rozróżnić między przynajmniej bilionem zapachów |
|
Yong |
|
2014-03-25 |
| Godzina Ziemi czyli o celebrowaniu ciemności |
|
Lomborg |
|
2014-03-27 |
| Słonie słyszą więcej niż ludzie |
|
Yong |
|
2014-03-30 |
| Niebo gwiaździste nade mną, małpa włochata we mnie |
|
Koraszewski |
|
2014-03-31 |
| Wielkoskrzydłe |
|
Naskręcki |
|
2014-04-02 |
| Najstarsze żyjące organizmy |
|
Coyne |
|
2014-04-03 |
| Jak zmienić bakterie jelitowe w dziennikarzy |
|
Yong |
|
2014-04-06 |
| Eureka! Sprytne wrony to odkryły |
|
Coyne |
|
2014-04-07 |
| Sukces upraw GM w Indiach |
|
Lomborg |
|
2014-04-09 |
| Wirus, który sterylizuje owady, ale je pobudza |
|
Yong |
|
2014-04-12 |
| Przystosować się do zmiany klimatu |
|
Ridley |
|
2014-04-14 |
| Jeden oddech, który zmienił planetę |
|
Naskręcki |
|
2014-04-16 |
| Najgorsze w karmieniu komarów jest czekanie |
|
Yong |
|
2014-04-17 |
| Kłopotliwa podróż w przyszłość |
|
Ridley |
|
2014-04-19 |
| Pierwsze spojrzenie na mikroby współczesnych łowców zbieraczy |
|
|
|
2014-04-23 |
| Seksizm w nauce o jaskiniowych owadach |
|
Coyne |
|
2014-04-26 |
| Musza bakteria zaprasza inne muszki na uczty owocowe |
|
Yong |
|
2014-04-27 |
| Zachwycający rabuś, który liczy sto milionów lat |
|
Cobb |
|
2014-04-28 |
| Mądrość (małych) tłumów |
|
Zimmer |
|
2014-04-29 |
| Tak bada się ewolucję inteligencji u zwierząt |
|
Yong |
|
2014-05-02 |
| Fantastyczna mimikra tropikalnego pnącza |
|
Coyne |
|
2014-05-03 |
| Dlaczego większość zasobównie wyczerpuje się |
|
Ridley |
|
2014-05-04 |
| Pomidory tworzą pestycydy z zapachu swoich sąsiadów |
|
Yong |
|
2014-05-07 |
| Potrawy z pasożytów |
|
Zimmer |
|
2014-05-08 |
| Technologia jest często matką nauki, a nie odwrotnie |
|
Ridley |
|
2014-05-09 |
| Montezuma i jego flirty |
|
Coyne |
|
2014-05-11 |
| Insekt dziedziczy mikroby z plemnika taty |
|
Yong |
|
2014-05-12 |
| Polowanie na nietoperze |
|
Naskręcki |
|
2014-05-14 |
| Zmień swoje geny przez zmianę swojego życia |
|
Coyne |
|
2014-05-15 |
| Obrona śmieciowego DNA |
|
Zimmer |
|
2014-05-17 |
| Gdzie są badania zwierzęcych wagin? |
|
Yong |
|
2014-05-20 |
| Niemal ssaki |
|
Naskręcki |
|
2014-05-21 |
| Zobaczyć jak splątane są gałęzie drzewa |
|
Zimmer |
|
2014-05-23 |
| Dlaczego ramiona ośmiornicy nie plączą się |
|
Yong |
|
2014-05-24 |
| Niezwykły pasikonik szklany |
|
Naskręcki |
|
2014-05-27 |
| Wąż zgubiony i ponownie odnaleziony |
|
Mayer |
|
2014-05-28 |
| Niespodziewani krewni mamutaków |
|
Yong |
|
2014-05-30 |
| Trochę lepszy świat |
|
Ridley |
|
2014-05-31 |
| Tam, gdzie są ptaki |
|
Mayer |
|
2014-06-01 |
| Ewolucja, ptaki i kwiaty |
|
Coyne |
|
2014-06-02 |
| Jestem spełniony |
|
Naskręcki |
|
2014-06-04 |
|
| « Poprzednia strona Następna strona » |