Alfred Sturtevant: bohater genetyki

W “Genetics” jest nowy artykuł Mariany Wolfner i Danny’ego Millera z ślicznym tytułem (poniżej), który naświetla inne osiągnięcia Sturtevanta: znalezienie nierównych crossing over między chromosomami. (Kliknij na link do artykułu: http://www.genetics.org/content/204/3/833 ).

O co chodzi z Sturtevantem wchodzącym do baru? No cóż, Sturtevanta zafrapowała obserwacja mutacji “Bar’ na chromosomie X u D. melanogaster, która powoduje małe oczy (patrz diagram poniżej). Muchy z oczyma Bar wykazywały niezwykle wysokie poziomy „mutacji”: około 1 na 1000 potomków samicy nosicielki tego genu albo powracało do “dzikiego typu” (normalne oczy), albo stawało się “ultra-Bar” (wyjątkowo wąskie oczy; patrz diagram dla obu).
To tempo mutacji było znacznie wyższe niż normalna mutacja genów (około 0,000001) i Sturtevant, opierając się na poprzednich swoich badaniach nad crossing-over między chromosomami (wymiana materiału genetycznego między parami “homologicznych” chromosomów podczas tworzenia się gamet), postawił hipotezę, że “mutacje” Bar nie były przypadkami zmian w samym genie, ale zmianami w budowie chromosomu wokół tego genu. Przez skomplikowaną serię krzyżówek genetycznych z użyciem zmutowanych genów otaczających odcinek Bar, Sturtevant był w stanie pokazać, że mutanty Bar powstają ze zjawiska znanego jako „nierówne crossing over”.
Normalnie podczas “mejozy”, procesu genetycznego tworzenia gamet w organizmach diploidalnych, “chromosomy homologiczne” zestawiają się w pary (mamy dwie kopie każdego z naszych chromosomów, a więc mamy 23 pary lub 46 razem; muchy Sturtevanta miały 8 razem). To zestawianie się w pary jest zasadnicze, by zapewnić, że homologi rozdzielają się, ponieważ każdy pójdzie do innego jaja lub plemnika (jaja i plemniki mają tylko połowę liczby chromosomów normalnych komórek i kiedy zachodzi zapłodnienie, powraca normalna liczba). Podczas zestawiania się w pary chromosomy homologiczne mogą wymieniać geny, kiedy rozłamują się i odcinki różnych homologów zlewają się z drugim homologiem. (Z jakiegoś powodu, który nie w pełni rozumiemy, samice Drosophila przechodzą ten proces, ale samce nie przechodzą. Właśnie obserwacja, że zmiany Bar widać było tylko od zmutowanych matek, ale nie ojców, która doprowadziła Sturtevanta do przypuszczenia, że chodzi o crossing-over zamiast o zwykłą mutację.)
Zazwyczaj crossing over jest dokładne z nukleotydami rozłamującymi się i zlewającymi w tym samym miejscu, jeśli więc jeden chromosom ma allel A, a drugi allel A’genu, zamienią się pozycjami w doskonałym porządku. (Oczywiście, reszta przylegających genów przejdzie razem.) Czasami jednak rekombinacja nie będzie doskonała i możesz otrzymać dwie kopie genu na jednym chromosomie i żadnej na drugim. Na przykład, ———A——— zestawione w parę z ———A’——— może dać ———AA’——— na jednym chromosomie i —————— na drugim. Jeden chromosom kończy z dwiema kopiami całego genu; drugi nie ma żadnej.
Przez używanie chytrych krzyżówek z mutacjami otaczającymi odcinek Bar, Sturtevant pokazał, że właśnie to powodowało zjawisko Bar. Kiedy muchy o normalnych oczach podlegały nierównemu crossing over, mogły dawać muchę z dwiema całymi kopiami odcinka genu, powodując wąskie oczy „Bar”. Mogły także, skojarzone z normalną muchą, dawać muchę z trzema kopiami odcinku genu i jeszcze węższymi „ultra-Bar” oczyma. Kiedy mucha z oczyma Bar traciła jedną ze swoich kopii dzięki nierównemu crossing over, powracała do stanu normalnej muchy.
W czasie, kiedy Sturtevant wykonywał te eksperymenty w latach 1920., nie było sposobu na potwierdzenie jego hipotezy przez bezpośrednie spojrzenie na chromosomy. Wkrótce potem jednak odkryto, że w gruczołach ślinowych much były „chromosomy poligeniczne”, w których DNA replikował się setki razy, a więc można było rzeczywiście obejrzeć fizyczną budowę chromosomów pod mikroskopem. Tutaj, na przykład, są dwa ramiona drugiego chromosomu, oba na fotografiach i w interpretacji. Fizyczne markery („paski”) chromosomów poligenicznych są diagnostyczne: te same dla wszystkich osobników gatunku.
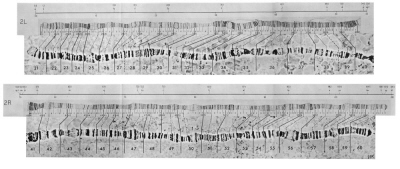
Kiedy Bridges (1936) i Muller et al. (1936) zbadali chromosomy z gruczołów ślinowych u much normalnych, much Bar i much ultra-Bar, potwierdzili, że oczy “Bar” istotnie pochodzą z duplikacji odcinka genu badanego przez Sturtevanta dziesięć lat wcześniej i że muchy ultra-Bar pochodzą z potrojenia tego odcinka, jak to pokazuje diagram poniżej z artykułu Wolfner i Millera.
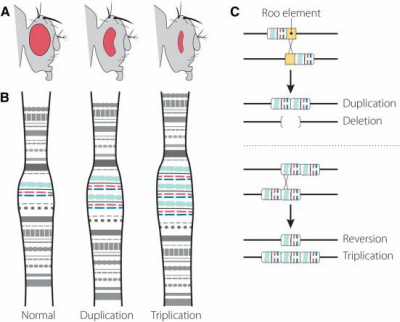
Dlaczego jest to ważne? Ponieważ nierówne crossing over jest jednym z głównych źródeł powstawania nowych genów w ewolucji. Prowadzi do duplikacji pojedynczego genu na jednym chromosomie, a kiedy to dzieje się, ewolucja może doprowadzić te dwie kopie do rozejścia się i przyjęcia nowych funkcji. Ponadto, nierówne crossing-over może stworzyć całe rodziny genów, które pochodzą od jednej kopii, ale ich funkcje rozeszły się po duplikacji, potrojeniu i tak dalej. Jest to jeden ze sposobów, na jakie genom rozwija się i na jaki możemy zyskać nowe geny z nowymi funkcjami. W ten sposób, na przykład, różne hemoglobiny: α, β, γ i δ, każda z inna funkcją, pochodzą od wspólnego przodka.
Oryginalne badanie Sturtevanta było więc zwiastunem naszego zrozumienia, jak powstaje nowa informacja genetyczna – przez pomyłkę w rekombinacji. Podobnie, nowa informacja genetyczna może powstać poprzez mutacje w jednym genie – także „pomyłce” w replikacji genu. Gdyby crossing over i replikacja genów zachodziły w sposób doskonały, nie byłoby ewolucji.
Sturtevant, nazywany przez wielu przyjaciół “Sturt”, był podobno wspaniałym człowiekiem, wolnym od obłudy i arogancji i (jak całe potomstwo Morgana – może z wyjątkiem H. J. Mullera) orzeźwiająco wolny od pragnienia zdobywania zasług za każde osiągnięcie. Żal mi, że nigdy nie spotkałem Sturtevanta, ale znam kilka osób, które znały go dobrze i bez wyjątku wszyscy mówią o nim jako o wielkim facecie.
Sturtevant został później profesorem genetyki w Cal Tech w Pasadenie, gdzie pozostał na resztę życia. I, jak porządny Drosophilista, do samego końca własnymi rekami popychał muszki. Tutaj jest jako już starszy mężczyzna we własnym „pokoju much”, robiąc coś, co jest obecnie zabronione w laboratoriach: paląc tuż obok pojemnika na eter, który usypia muchy i jest WYSOCE ŁATWOPALNY.

h/t: Matthew Cobb
Alfred Sturtevant a hero of genetics
Why Evolution Is True, 24 stycznia 2017
Tłumaczenie: Małgorzata Koraszewska

Profesor (emeritus) na wydziale ekologii i ewolucji University of Chicago, jego książka "Why Evolution is True" (Polskie wydanie: "Ewolucja jest faktem", Prószyński i Ska, 2009r.) została przełożona na kilkanaście języków, a przez Richarda Dawkinsa jest oceniana jako najlepsza książka o ewolucji. Jerry Coyne jest jednym z najlepszych na świecie specjalistów od specjacji, rozdzielania się gatunków. Jest wielkim miłośnikiem kotów i osobistym przyjacielem redaktor naczelnej.