Depresja inbredowa u człowieka
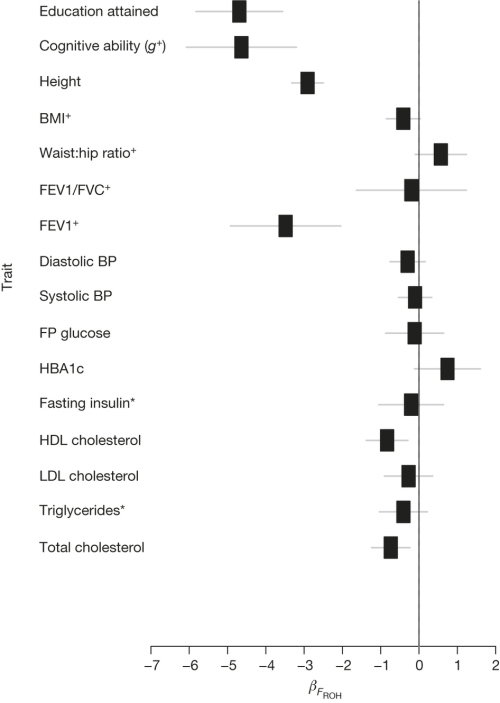
Jest to ciekawy, ale spodziewany, wynik. Od dawna było wiadomo, że kojarzenie się bliskich krewnych zarówno u roślin, jak u zwierząt, może prowadzić do zmniejszonej żywotności, zmniejszonego wigoru i wad fenotypu: depresji inbredowej. Chociaż hodowcy od dawna znali to zjawisko, pierwszy zbadał je Darwin (który martwił się możliwymi skutkami tego u własnych dzieci, jako że jego żona, Emma, była jego kuzynką pierwszego stopnia); badał on skutki kojarzenia krewniaczego i kojarzenia niekrewniaczego u wielu roślin, z których znaczna część miała adaptacje ograniczające rozmiary inbredu i zapewniające kojarzenie z niekrewnymi. Często mówi się także, że problem inbredu dotykał rodzin królewskich w Europie, które wielokrotnie zawierały małżeństwa w małej grupie rodzin. Słynny w historii przypadek, przypisywany kojarzeniu krewniaczemu, dotyczy “wargi habsburskiej”, nie był jednak spowodowanie kojarzeniem krewniaczym, ponieważ allel powodujący prognatyzm jest dominujący (patrz poniżej, dlaczego jest to istotne), chociaż inbred mógł przyczynić się do fizycznego i psychicznego upadku tej rodziny.

Odwrotność depresji inbredowej – wigor hybryd – także był znany od dawna: potomstwo niespokrewnionych osobników lub różnych szczepów tego samego gatunku wykazuje często wzmożony wigor, wzmożoną żywotność i wzmożoną płodność. Niemal cała kukurydza rosnąca na farmach w Stanach Zjednoczonych pochodzi z nasion produkowanych przez krzyżowanie różnych odmian. Tak zwana „kukurydza hybrydowa” daje wyższe plony niż odmiany rodzicielskie (a także zapewnia, że firmy nasiennicze otrzymują zapłatę corocznie, ponieważ odmiana wysokoplenna nie może odrodzić się następnego roku, jeśli farmer zachowuje trochę tegorocznych plonów na nasiona). Podobne zjawisko może zachodzić w krzyżówkach międzygatunkowych, ale potomstwo takich skrzyżowań, mimo że jest duże i z dużym wigorem, może być bezpłodne (np. muły, skrzyżowanie koni i osłów) tak więc o takich sterylnych krzyżówkach mówi się, że wykazują bujność somatyczną.
W genetyce od dawna trwa debata o przyczynach wigoru hybryd/depresji inbredowej. Są dwie główne możliwości. Pierwsza, osobniki inbredowe mają zredukowany wigor, ponieważ jest bardziej prawdopodobne, że są homozygotyczne (tj. posiadają dwie kopie) szkodliwych mutacji recesywnych. U heterozygoty szkodliwe efekty allelu recesywnego są maskowane przez allel dominujący, podczas gdy u homozygoty takie szkodliwe efekty mogą być wyrażane. Ponieważ zaś głównym skutkiem genetycznym krzyżowania krewniaczego jest podniesienie homozygotyczności, występują fenotypowe efekty szkodliwych alleli recesywnych. Drugą możliwością jest to, że osobniki inbredowe mają mniejsze prawdopodobieństwo heterozygotyczności w genach, które wykazują naddominację dostosowania, a więc wyrazi się to w mniej dostosowanych fenotypach związanych z homozygotycznymi genotypami. W naddominacji dostosowania heterozygoty mają największą żywotność i/lub płodność, podczas gdy obie homozygoty mają niższą. Najbardziej chyba znanym przykładem naddominacji dostosowania jest allel anemii sierpowatej w ludzkiej hemoglobinie w środowiskach malarycznych: heterozygoty nie chorują na anemię sierpowatą oraz są odporne na malarię, uzyskując w ten sposób wyższe dostosowanie niż każda z homozygot. (W niemalarycznym środowisku dostosowanie heterozygot jest zasadniczo takie samo jak dzikiego typu homozygot.) Oba te zjawiska genetyczne – szkodliwe geny recesywne i naddominacja dostosowania – mogą prowadzić do depresji inbredowej. W obszernym przeglądzie literatury kilka lat temu Deborah Charlesworth i John Willis pokazali, że główną przyczyną są szkodliwe geny recesywne, a naddominacja przykłada się do tego, ale jej wkład jest niewielki.
Stosunek między dominacją a dostosowaniem figuruje także w innej długotrwałej debacie w genetyce ewolucyjnej: debacie między R.A. Fisherem a Sewallem Wrightem, dwoma założycielami teoretycznej genetyki populacji, o to, czy nowe, szkodliwe mutacje są recesywne ab initio (fide Wright), czy też nacisk selekcyjny na modyfikowanie alleli powoduje, że początkowo dominujące lub addytywne działania szkodliwych mutacji stają się recesywne (fide Fisher). Wright pokazał, że selektywne skutki takich modyfikatorów byłyby bardzo małe (mniej więcej tej samej mocy, co tempo mutacji, które jest bardzo małe) i wątpił, by taka skromna selekcja mogła przeważyć nad innymi czynnikami (włącznie z doborem innych skutków fenotypowych modyfikowanych alleli) w populacjach naturalnych. Fisher, który uważał, że populacje naturalne są duże, sądził, że mogła. Fakt, że nowo zaobserwowane mutacje są na ogół recesywne i pomysłowa praca studenta Jerry’ego Coyne’a, Allena Orra, który użył normalnie haploidalnych glonów, żeby pokazać, iż recesywność jest regułą także kiedy nie ma możliwości doboru modyfikujących alleli w stanie diploidalnym, przekonały wreszcie większość ludzi, “że recesywne efekty fenotypowe rzadkich mutacji nie wynikają z selekcji dominujących modyfikatorów”. (Charlesworth and Charlesworth, 2010:183).
Badanie Joshi’ego skupia się na stosunku cech fenotypowych i inbredzie wewnątrz populacji, nie mówi więc niczego bezpośrednio o skutkach małżeństw między grupami etnicznymi i narodowymi. Dla na ogół addytywnych, poligenetycznych cech, takich jak wzrost, oczekuje się, że dzieci takich małżeństw miałyby wzrost pośredni wobec obojga rodziców (nie wyższy niż oboje), ale nie można wykluczyć wigoru hybryd w innych cechach.
Alvarez, G., F.C. Ceballos and C. Quintero. 2009. The role of inbreeding in the extinction of a European royal dynasty. Plosone 4(4): e5174, 7 pp. pdf
Charlesworth, B. and D. Charlesworth. 2010. Elements of Evolutionary Genetics. Roberts, Greenwood Village, Colorado. (pp. 170-183)
Charlesworth, D. and J.H. Willis. 2009. The genetics of inbreeding depression. Nature Reviews Genetics 10:783-796. pdf
Darwin, C. 1876. The Effects of Cross and Self Fertilisation in the Vegetable Kingdom. John Murray, London. (Darwin Online)
Joshi, P.K. et al. 2015. Directional dominance on stature and cognition in diverse human populations. Nature in press. html
Orr, H. A. 1991. A test of Fisher’s theory of dominance. Proceedings of the National Academy of Sciences USA 88: 11413-11415. pdf
Provine, W.B. 1986. Sewall Wright and Evolutionary Biology. University of Chicago Press, Chicago. (pp. 243-260)
Thompson, E.M. and R.M. Winter. 1988. Another family with the ‘Habsburg jaw’. Journal of Medical Genetics 25: 838-842. pdf
Why Evolution Is True, 3 lipca 2015
Tłumaczenie: Małgorzata Koraszewska

Greg Mayer
Zastępca kuratora działu gadów i płazów w muzeum zoologicznym Uniwersytetu Wisconsin. Doktorat z biologii ewolucyjnej obronił na Harvardzie.