

Wzrastająca kolonia Escherichia coli (Wikipedia)
Gatunek biologiczny składa się z grupy populacji, które tam, gdzie współistnieją w przyrodzie, wymieniają geny poprzez rozmnażanie. Dwie populacje, które współistnieją na jednym obszarze, ale nie wymieniają genów, są uważane za członków różnych gatunków biologicznych.
Jedną z zalet BSC jest to, że umożliwia nam natychmiastowe rozwiązanie problemu gatunku, który umykał Darwinowi: dlaczego przyroda jest podzielona na odrębne zgrupowania, a nie istnieje jako kontinuum, zgrupowania najbardziej widoczne tam, gdzie współistnieją? Zgodnie z BSC problem „pochodzenia gatunków” staje się po prostu problemem „pochodzenia barier uniemożliwiających krzyżowanie się” – i jest to problem możliwy do rozwiązania. (Ponownie, zobacz Coyne'a i Orra, aby poznać nasze podejście do powstawania tych zgrupowań.)
Oczywiście są problemy z tym pojęciem (nie jest to definicja aprioryczna, ale próba konceptualizacji słowami tego, co widzimy w naturze). Problemy te obejmują ocenę populacji żyjących na różnych obszarach, takich jak wyspy archipelagu, sposób radzenia sobie z grupami, które w niewielkim stopniu tworzą hybrydy tam, gdzie współistnieją oraz, co najważniejsze w tym artykule, co robimy z gatunkami bezpłciowymi, pozbawionymi możliwości wymiany genów. Wszystkie te kwestie omawiamy w pierwszym rozdziale mojej książki Speciation (2009) napisanej z Allenem Orrem, ale jedną z kwestii, których nie rozwiązaliśmy właściwie, była kwestia organizmów bezpłciowych.
A co z tymi uprzykrzonymi organizmami „bezpłciowymi”? Jak możemy konceptualizować gatunki w grupach takich jak bakterie? Cóż, pierwszą rzeczą, jaką musimy ustalić, jest to, czy tworzą rozróżnialne zgrupowania, takie jak ptaki czy żółwie. Jeśli tak nie jest, nie ma potrzeby konceptualizowania nieistniejących zgrupowań. W naszej książce z 2009 roku dokonaliśmy przeglądu literatury, która była wówczas skąpa, i zdecydowaliśmy, że dowody na to, czy bakterie (uważane za bezpłciowe) tworzą gatunki, były mieszane, ale z pewnością były wśród nich pewne zgrupowania. Dlatego ograniczyliśmy resztę książki do organizmów rozmnażających się płciowo. Mimo to „gatunkom” bakterii nadaje się nazwy, takie jak E. coli, ale czy wszystkie bakterie uważane za E. coli naprawdę stanowią członków odrębnego zgrupowania? Jeśli tak, to jak?
Od tego czasu literatura się rozwinęła, a poniższy artykuł, który przeoczyłem i który ma już siedem lat, całkiem nieźle udowadnia, że przynajmniej wśród bakterii istnieją gatunki i, co ważniejsze, są one konceptualizowane w sposób podobny do BSC. Innymi słowy, istnieją zgrupowania bakterii, a każde zgrupowanie charakteryzuje się zdolnością do wymiany genów między osobnikami. Członkowie różnych zgrupowań nie wymieniają jednak genów. Innymi słowy, bakterie składają się głównie z genetycznie izolowanych zgrupowań. Autorzy, choć badają jedynie bakterie (istnieją inne organizmy rozmnażające się bezpłciowo, jak wrotki bdelloidalne), dochodzą do wniosku, że życie w ogóle jest zgodne z BSC. To trochę zbyt ekspansywny wniosek (patrz tytuł!), ale ich wyniki dla bakterii wydają się dobre.
Kliknij na link, aby przeczytać, lub zobacz plik PDF tutaj.
Kluczem do tego artykułu jest uznanie, że bakterie w rzeczywistości nie są całkowicie bezpłciowe, chociaż często rozmnażają się w ten sposób. Ale mają też taką formę seksu, w której genomy dwóch różnych osobników mogą zbliżyć się do siebie i rekombinować, tworząc nowe geny. Proces ten, zwany rekombinacją homologiczną, zachodzi poprzez kontakt między komórkami lub przeniesienie DNA przez rurki („pilusy”) łączące różne jednostki. Proces ten nazywa się koniugacją.
Oto zdjęcie z Wikipedii przedstawiające dwie komórki bakteryjne przemieszczające DNA przez pilusy:
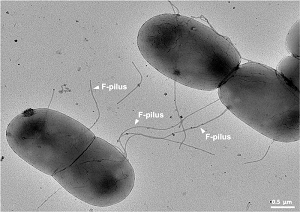
Ten plik udostępniony jest na licencji Creative Commons Attribution-Share Alike 4.0 International.
Ten ruch jest jednokierunkowy: DNA (pojedynczy chromosom z dwuniciowym DNA) od jednego osobnika przemieszcza się do drugiego. Następnie może nastąpić forma rozmnażania „płciowego”, w której różne kopie tego samego genu mogą ustawić się obok siebie i rekombinować, tworząc nowy gen. Podobny proces zachodzi podczas mejozy (tworzenia gamet) w organizmach rozmnażających się płciowo.
U bakterii to mieszanie się podobnych genów nazywa się rekombinacją homologiczną, ponieważ zmienia skład genu poprzez rekombinację jego DNA z DNA podobnego genu w innej bakterii. Istnieją inne formy wymiany DNA u bakterii, w których fragment DNA lub „plazmid” jednego osobnika po prostu wstawia się gdzieś indziej w genomie innego osobnika, ale nie jest to rekombinacja w tradycyjnym sensie, ponieważ nie obejmuje dwóch różnych kopii tego samego genu, które dokonują rekombinacji w celu utworzenia nowego genu. Artykuł Bobay i Ochmana dotyczy rekombinacji homologicznej.
Stosowana przez nich metoda określania, czy dwa osobniki danego gatunku bakterii mogą w ten sposób rekombinować swoje DNA, jest skomplikowana i lepiej pozostawię ją ekspertom. Powiem jednak, że polega to na wykazaniu, że jednostki w grupie mają te same warianty w danym segmencie genu (zsekwencjonowano 10 000 zasad), jak inne jednostki w grupie. Na przykład, w jednym dziesięciozasadowym odcinku DNA, osobnik może mieć GTTACTCTAA, inny będzie miał GTTAGTCTAA, jeszcze inny GTTACTCTAC, a jeszcze inny GTTACTAC, reprezentujący kombinacje zasad DNA, które mogą wystąpić w wyniku rekombinacji.
Jeśli widzisz ten wzór wśród osobników określonego gatunku bakterii, oznacza to, że ma miejsce rekombinacja homologiczna – bakteryjny „seks” zachodzi. Ta forma rekombinacji nazywana jest rekombinacją „homoplastyczną”, ponieważ wszystkie warianty powstają w wyniku mutacji pojedynczego pierwotnego genomu obecnego u osobnika, który założył gatunek.
Jedną z alternatyw jest to, że mamy do czynienia z dwoma spokrewnionymi gatunkami, u których podobne sekwencje DNA wyglądają jedynie tak, jakby przeszły rekombinację homologiczną, ponieważ dwie grupy miały wspólnego przodka, a następnie potomkowie mieli podobne („zbieżne”) mutacje. Zjawisko to, zwane „rekombinacją niehomoplastyczną”, nie jest spowodowane wymianą genetyczną.
Autorzy mają sposoby na rozróżnienie tych dwóch typów rekombinacji i opracowali stosunek, który nazywają „h/m”, pokazujący stosunek stopnia rekombinacji homoplastycznej (prawdziwy „seks”) do rekombinacji niehomoplastycznej (niezależne mutacje w różnych grupach, które powierzchownie naśladują „seks” ). Im wyższy stosunek h/m, tym więcej osobników w tej grupie uprawia „seks”.
Autorzy obliczyli stosunki h/m dla 91 nazwanych „gatunków” bakterii, używając oczywiście dużej liczby sekwencji genomów dla każdego gatunku, ponieważ trzeba zbadać zmienność między osobnikami w tym segmencie liczącym 10 000 zasad. (Przeprowadzili także symulacje, aby sprawdzić, czy potrafią odróżnić „h” od „m”.) Okazuje się, że ponad połowa z 91 nazwanych gatunków bakterii, które zbadali, pasowała do gatunków biologicznych, w przypadku których istniały dowody na rekombinację „h” między osobnikami. Oto jeden poniżej, w którym wzrasta stosunek h/m osiągając asymptotę, gdy przyglądali się większej liczbie szczepów. (Zwiększa to zdolność wykrywania udostępnionych wariantów). 54 z 91 nazwanych gatunków bakterii tak wyglądało, więc BSC obejmuje co najmniej połowę nazwanych gatunków bakterii, a autorzy pobrali duże próbki bakterii.
Gatunek biologiczny u bakterii:
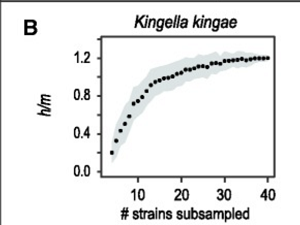
Tutaj można zobaczyć to, co uznano za jeden gatunek, ale po dodaniu większej liczby szczepów zaobserwowano dwa zgrupowania, z których jedno zachowywało się jak powyżej, ale drugie, w porównaniu z pierwszym, wykazywało bardzo niski stosunek h/m, co wskazuje, że między tymi grupami nie występowała rekombinacja homologiczna. Oznacza to, że były to różne „gatunki biologiczne”. Kiedy wyeliminowano grupę o niskim h/m, B. pseudomallei zachował się jak trzeba. Zatem mamy tu dwa gatunki, którym nadano tę samą nazwę, być może dlatego, że miały podobną morfologię lub wymagania hodowlane, albo dlatego, że odległość genetyczna między nimi (wskazująca na czas separacji) była dość mała, co sugeruje niedawne pochodzenie. Te „tajemnicze gatunki” zaobserwowano u 21 z 91 wymienionych gatunków bakterii.
Dwa gatunki biologiczne bakterii, które występowały pod jedną nazwą:

A trzecia grupa sama w sobie miała niski stosunek h/m, niezależnie od tego, ile szczepów obejmowała, więc w ogóle nie było możliwości oceny przepływu genów – być może dlatego, że te gatunki po prostu nie przechodzą żadnej rekombinacji homologicznej. Tu jest jeden:
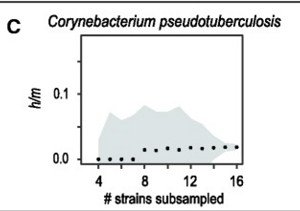
Zatem 73 z 91 przebadanych grup wykazało wzorce zgodne z koncepcją gatunku opartą na izolacji reprodukcyjnej.
Aby sprawdzić, czy ich metoda rzeczywiście wykryła grupy analogiczne do gatunków biologicznych u bardziej znanych zwierząt, autorzy przeprowadzili ten sam rodzaj testu h/m dla dwóch par spokrewnionych, ale wyraźnie odrębnych gatunków biologicznych; jednym z nich był pokrewny gatunek Drosophila melanogaster i D. simulans, a drugim Homo sapiens i szympans Pan troglodytes. Jak widać poniżej, udało im się wykryć izolację reprodukcyjną pomiędzy grupami przy użyciu podobnego fragmentu o długości 10 000 par zasad. (We wszystkich przypadkach porównywano wiele tych samych sekwencji różnych osobników po lewej stronie z jedną sekwencję gatunku po prawej, dlatego „inny” gatunek tworzy linię prostą: mamy jedną sekwencję w porównaniu z wieloma sekwencjami w drugim gatunków, a wszystkie porównania wykazują niski stosunek h/m.)
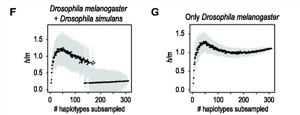
Ludzie kontra szympansy, także dobre gatunki biologiczne:
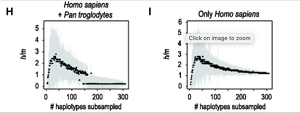
Wniosek jest zatem taki, że BSC całkiem nieźle radzi sobie z konceptualizacją gatunków bakterii: istnieją grupy, które wymieniają segmenty genów i inne grupy (różne „gatunki”), które nie wymieniają DNA poprzez rekombinację homologiczną. Pamiętajcie: wszystko to zostało ocenione na podstawie sekwencji DNA, a nie bezpośredniej wymiany genów.
Najważniejszy wniosek (z artykułu):
Fakt, że gatunek można uniwersalnie zdefiniować na podstawie przepływu genów, oznacza, że w procesie specjacji we wszystkich formach życia działa wiele tych samych czynników. Różnice we właściwościach genomicznych (takich jak ploidalność, częstotliwość rekombinacji i rozmnażanie oraz tempo nabywania genów) i w parametrach demograficznych (takich jak wielkość populacji, rozmieszczenie geograficzne i tempo migracji) będą miały wpływ na tempo specjacji drobnoustrojów w stosunku do organizmów płciowych. Jednakże zastosowanie pojedynczego kryterium BSC opartego na genomie do wytyczenia gatunków umożliwia zdefiniowanie gatunków i badanie specjacji w podobnych ramach w całym drzewie życia.
Cóż, muszą przyjrzeć się innym domniemanie bezpłciowym grupom, aby sprawdzić, czy ta metoda pokazuje również istnienie krzyżujących się grup reprodukcyjnie izolowanych od innych tego typu grup, ale przynajmniej w przypadku bakterii widzimy, że wiele z nich tworzy zgrupowania. Pozostają dwa pytania:
1.) Czym zatem jest „specjacja” u bakterii? Jednym z najbardziej intrygujących wyników tej pracy jest to, że jeśli weźmie się pod uwagę pary „gatunków” bakterii, stopień izolacji reprodukcyjnej między nimi nie jest dodatnio skorelowany z czasem, jaki je dzieli, ocenianym na podstawie „odległości genetycznej” lub całego genomu rozbieżność między nimi. Trudno to zrozumieć, ponieważ sugeruje, że w przeciwieństwie do organizmów rozmnażających się płciowo, takich jak muszki owocowe i ssaki, bariery reprodukcyjne nie tworzą się jako prosty produkt uboczny czasu oddzielenia się od wspólnego przodka. Dzieje się tak, ponieważ w tych grupach bariery reprodukcyjne są zwykle produktem ubocznym rozbieżności między populacjami w wyniku doboru naturalnego i dryfu genetycznego, które z biegiem czasu genetycznie oddzielają gatunki.
Dlaczego nie dzieje się tak w przypadku bakterii? Nie mam pojęcia! Moją jedyną sugestią jest to, że „tworzenie się gatunku” może zachodzić u bakterii tak szybko, że po prostu nie da się korelować czasu z izolacją reprodukcyjną. Oznaczałoby to, że jest to praktycznie natychmiastowe.
2.) Dlaczego bakterie tworzą zgrupowania? U bardziej znanych zwierząt zgrupowania powstają, ponieważ po powstaniu barier reprodukcyjnych grupa krzyżująca się może swobodnie przystosować się do swojego środowiska bez „zanieczyszczania” przez inne gatunki, które mogłoby rozmyć zgrupowanie. Dywergencja genetyczna znajduje odzwierciedlenie nie tylko w izolacji reprodukcyjnej, ale także w wyglądzie i zachowaniu organizmów. Może to dotyczyć również bakterii: każdy klaster może reprezentować grupę przystosowaną do określonej niszy ekologicznej. Trudno byłoby to przetestować na bakteriach występujących w naturze, ale można by to przetestować na bakteriach chorobotwórczych, których siedlisko (nas) można łatwiej zbadać. O ile pamiętam, każdy gatunek bakterii działa na swój sposób, ale to nie jest odpowiedź na pytanie.
_______________
Reference: Bobay LM, Ochman H. Biological species are universal across Life’s domains. Genome Biol Evol. 2017 Feb 10;9(3):491–501. doi: 10.1093/gbe/evx026.
Link do oryginału: https://whyevolutionistrue.com/2024/02/25/do-asexual-bacteria-form-biological-species/
Why Evolution Is True, 25 lutego 2024
Tłumaczenie: Małgorzata Koraszewska

Jerry A. Coyne
Emerytowany profesor na wydziale ekologii i ewolucji University of Chicago, jego książka "Why Evolution is True" (Polskie wydanie: "Ewolucja jest faktem", Prószyński i Ska, 2009r.) została przełożona na kilkanaście języków, a przez Richarda Dawkinsa jest oceniana jako najlepsza książka o ewolucji. Jerry Coyne jest jednym z najlepszych na świecie specjalistów od specjacji, rozdzielania się gatunków. Jest również jednym ze znanych "nowych ateistów" i autorem książki "Faith vs Fakt" (wydanej również po polsku przez wydawnictwo "Stapis)". Jest wielkim miłośnikiem kotów i osobistym przyjacielem redaktor naczelnej.
| Chief editor: | Hili |
| Webmaster:: | Andrzej Koraszewski |
| Collaborators: | Jacek Chudziński, Hili, Małgorzata Koraszewska, Andrzej Koraszewski, Henryk Rubinstein |
