“Partenogenetyczny” rak rozmnaża się bez seksu: czy to jest nowy gatunek?

Spójrzmy jednak na oryginalne prace (linki poniżej), żeby zobaczyć, co się dzieje.
Można to streścić dość prosto, choć genetyka jest skomplikowana. W 1995 r. “rak marmurkowy” pojawił się w niemieckim handlu zwierząt do akwarium, pochodząc prawdopodobnie od jednego mutanta, który był członkiem gatunki Procambarus Falla. Oto jak wygląda rak marmurkowy z pancerzem w plamki, podobnie jak jego bezpośredni przodek, P. fallax:

Jest to jedyny znany partenogenetyczny gatunek dziesięcionogów (grupa ta obejmuje krewetki, raki, homary i kraby); i rozmnaża się przez apomiktyczną partenogenezę – jajo tworzy się przez normalny podział komórkowy czyli mitozę i jest płodne. Żaden plemnik nie jest potrzebny! To daje jednemu osobnikowi zdolność zajęcia habitatu i zaludnienia go, co jest możliwe w płciowo rozmnażających się organizmach tylko w wypadku, kiedy samica jest zapłodniona.
Partenogeneza w połączeniu z wszystkożernością raka marmurkowego oznacza, że jest to zagrożenie i to zagrożenie, które się szerzy, szczególnie, jak pokazuje praca Gutekunst i in., na Madagaskarze. Partenogeneza niekoniecznie oznacza, że gatunek (w rzeczywistości jest to klon, jak zobaczymy za chwilę) zastąpi inne raki. Może być ekologicznie inne nie tylko od innych gatunków raka, ale od swoich przodków P. fallax, a w takim wypadku może żyć obok nich. Według teorii genetyki populacyjnej oczekuje się, że “mutanty” takie jak ten, zastąpią formy płciowe w tym samym gatunku, ale tylko jeśli ich ekologia jest taka sama. Niemniej zdolność tych stworzeń do jedzenia wszystkiego i do inwazji na jeziora słodkowodne przez jednego tylko osobnika jest niepokojącym zagrożeniem.
Jak to powstało? Jej powstanie jest w istocie kluczem do jej zdolności rozmnażania się bezpłciowego. Jest to triploid: zamiast normalnego zestawu 184 chromosomów (dwóch zestawów po 92) ma trzy zestawy: 276. (To bardzo dużo chromosomów – ludzie mają tylko 46). Prawdopodobnie P. fallax stworzył diploidalne jajeczko z 184 chromosomami zamiast normalnych 92. Potem doszedł do jajeczka plemnik z 82 chromosomami od innego osobnika, co dało triploid z 276 chromosomami.
Triploid nie może tworzyć zdolnych do życia osobników przez normalną mejozę, bo byłyby to trzy chromosomy każdego typu, starające połączyć się w pary, a potem rozdzielić, co dałoby bałagan. Jeśli w ten sposób w ogóle powstałoby jajeczko, to nie miałoby pełnego zestawu chromosomów, ale mogłoby mu brakować jednych, a innych byłby nadmiar, co przy zapłodnieniu dałoby zygotę o zupełnie zakłóconym rozwoju. Mogłoby jednak zreprodukować się przez normalny podział komórkowy (mitozę), bo to nie wymaga łączenia się chromosomów w pary. I w ten sposób rozmnaża się ten nowy klon: produkując jajeczka przez zwykły podział komórek somatycznych.
Musi także być jakaś cecha raka marmurkowego, która pozwala na rozwój triploidalnej komórki bez potrzeby zapłodnienia przez plemnik. (Te klony mogą kojarzyć się z samcami P. fallax, ale przekazane plemniki niczym nie przyczyniają się do potomstwa.) Nie wiemy, czy cecha, która pozwala temu klonowi na rozmnażanie bez seksu jest genetyczną mutacją, czy też po prostu jest to produkt uboczny komórki posiadającej trzy zestawy chromosomów.
Żeby spojrzeć na pochodzenie tego stworzenia, Gutekunst i in. zsekwencjonowali jego pełen genom: pierwszy w pełni zsekwencjonowany genom jakiegokolwiek skorupiaka dziesięcionoga. I znaleźli to, czego można było oczekiwać z powyżej przedstawionej teorii jego pochodzenia: osobnik miał trzy zestawy chromosomów, z których dwa były niemal identyczne, a trzeci nieco odbiegający, ale nadal był to genom P. fallax. To znaczy, że gatunek z pewnością powstał, jak przewidywano, z niezredukowanej komórki jajowej samicy P. fallax zapłodnionej plemnikiem samca P. fallax. Porównując DNA klona z innymi stawonogami, autorzy znaleźli tę filogenezę, pokazującą gatunek najbardziej zbliżony do krewnego raka, Parhyale hawaiensis, skorupiaka obunoga.
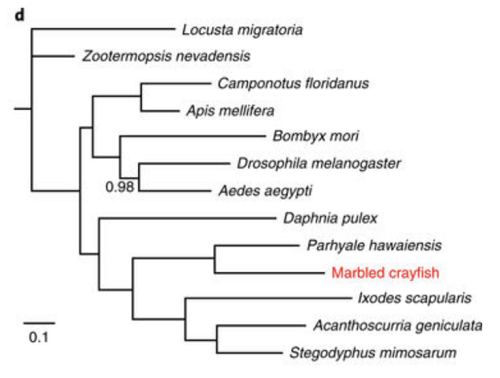
Mamy więc tu klona, który rozmnaża się bez seksu, podobnie jak te wirusy i bakterie, które reprodukują się bez prawdziwego, mejotycznego seksu (chociaż te mikroby mają sposób na wymianę genów).
Ale czy jest to nowy gatunek? Artykuł Franka Lyko (odnośnik poniżej) mówi, że jest i nazywa triploidalnego klona Procambarus virginalis. Na jakiej podstawie uznaje to za nowy gatunek? Nie jest nią morfologia, bo nie ma cechy, która absolutnie odróżnia tego triploidalnego klona od P. fallax. Genetycznie jednak można odróżnić klona od gatunku rodzicielskiego, ponieważ ma diagnostyczny mitochondrialny DNA. Niemniej jest tak tylko dlatego, że pierwszy zmutowany osobnik miał unikatową sekwencję DNA w mitochondriach (wszyscy mamy!), która została przekazana klonowym potomkom. Dla mnie jest to tylko arbitralna cecha, która umożliwia odróżnienie klonów od przodka, ale nie ma biologicznego znaczenia.
Lyko pisze, że klonowy “gatunek” jest izolowany reprodukcyjnie od P. fallax: nie mogą wymieniać genów. Izolacja reprodukcyjna jednej grupy od drugiej jest, oczywiście, biologicznym znakiem probierczym gatunku, zasługuje to więc na wzięcie pod uwagę. Ale samo to nie znaczy, że P. virginialis trzyma się klasycznej definicji biologicznego gatunku z tej przyczyny: osobniki w gatunku klonalnym są także izolowane reprodukcyjnie wzajem od siebie! Według biologicznej koncepcji gatunku (BSC) gatunkiem jest “grupa krzyżujących się ze sobą osobników, izolowanych reprodukcyjnie od innych takich grup”. Ale osobniki P. virginialis nie krzyżują się: nie mogą kojarzyć się wzajem ze sobą!
Tak więc pozostawione same w różnych jeziorach te izolowane klony mogą w nieskończoność rozchodzić się drogą ewolucji, a nikt nigdy nie będzie mógł sprawdzić ich zgodności reprodukcyjnej ani przez obserwacje w akwariach, ani przez zobaczenie, co dzieje się, kiedy znowu mają ze sobą kontakt. W obu wypadkach nie da się osądzić, czy mogą krzyżować się ze sobą.
Znaczy to, że podobnie jak bakterie lub inne grupy bezpłciowe, nie możemy użyć BSC, żeby stwierdzić, czy ten klon reprezentuje nowy gatunek. Wyznaczanie gatunków w takich grupach staje się mniej lub bardziej arbitralne i opiera się o stopnie genetycznych lub morfologicznych różnic. Nie daje się powiedzieć, czy populacje klonów pozostawionych samotnie każdy w innym jeziorze staną się różnymi gatunkami. Czy różnica jednego nukleotydu w olbrzymim genomie, charakterystyczna dla danego jeziora, wystarcza, do oddzielenia nowego gatunku w ramach P. virginialis? Jeśli nie, to jaka różnica jest niezbędna? Z tego powodu łatwiej stosować pojęcie gatunku do organizmów rozmnażających się płciowo: jeśli spotykają się w naturze, albo krzyżują się ze sobą, albo nie (mogą oczywiście być problematyczne przypadki bardzo ograniczonego krzyżowania, ale u wielu grup ten problem nie występuje).
Nazwanie tego przez Lyko nowym gatunkiem jest decyzją różniącą się zasadniczo od decyzji uznania Drosophila simulans za inny gatunek niż D. melanogaster, ponieważ występują razem w przyrodzie, ale nigdy nie wymieniają genów (hybrydy są bezpłodne i większość z nich nie nadaje się do życia).
Gatunkami nazywano inne partenogenetyczne zwierzęta, jak słynną rybę Poeciliopsis monacha-occidentalis, która powstała z hybrydyzacji między dwoma gatunkami, ale rozmnaża się bezpłciowo od ponad 100 tysięcy pokoleń. Ona także jest teraz klonem a określenie jej jako gatunku jest mniej lub bardziej arbitralne, jeśli weźmie się pod uwagę, że nic nie promuje krzyżowania się ani nie przeszkadza genetycznemu rozchodzeniu się grup klonów.
Taki więc jest mój pogląd na kwestię tego, czy ta rzecz jest nowym gatunkiem: jest to problematyczne, ponieważ ten rak jest aseksualnym klonem. Biologia jest jednak fascynująca i byłoby dobrze odkryć, jaka cecha oryginalnego, triploidalnego osobnika umożliwiła mu rozmnażanie się bez seksu.
____________
Gutekunst, J., R. Andriantsoa, C. Falckenhayn, K. Hanna, W. Stein, J. Rasamy, and F. Lyko. 2018. Clonal genome evolution and rapid invasive spread of the marbled crayfish. Nature Ecology & Evolution, online, doi:10.1038/s41559-018-0467-9
Lyko, F. 2017. The marbled crayfish (Decapoda: Cambaridae) represents an independent new species. Zootaxa 4363:544-552.
A “parthenogenetic” crayfish reproduces without sex: is it a new species?
Why Evolution Is True, 8 lutego 2018
Tłumaczenie: Małgorzata Koraszewska

Profesor (emeritus) na wydziale ekologii i ewolucji University of Chicago, jego książka "Why Evolution is True" (Polskie wydanie: "Ewolucja jest faktem", Prószyński i Ska, 2009r.) została przełożona na kilkanaście języków, a przez Richarda Dawkinsa jest oceniana jako najlepsza książka o ewolucji. Jerry Coyne jest jednym z najlepszych na świecie specjalistów od specjacji, rozdzielania się gatunków. Jest wielkim miłośnikiem kotów i osobistym przyjacielem redaktor naczelnej.