Specjacja przez hybrydyzację u amazońskich gorzyków?
Ale mamy nowe doniesienie o specjacji przez hybrydyzację u gorzyków z Amazonii i wygląda ono całkiem dobrze. Opublikowane w PNAS (odnośnik poniżej, pdf tutaj), pierwszym autorem jest Anfredo Barerra-Guzmán, a ostatnim Jason Weir, badacz po doktoracie z laboratorium Trevora Price’a, który pracował przez kilka lat drzwi w drzwi z moim.
Artykuł jest długi i skomplikowany z pewnymi genetycznym szczegółami, których niespecjaliści nie potrzebują, i streszczę wyniki tak zwięźle, jak potrafię.
W brazylijskiej Amazonii, w górnym biegu rzeki Cururu-ri w stanie Pará, żyją trzy gatunki gorzykowatych: Lepidothrix vilasboasi, L. nattereri i L. iris, który ma dwa podgatunki. L. vilasbosi, którego będę nazywał LV, żyje na terenie pomiędzy L. nattereri (“LN”) i L. iris (“LI”); wszystkie są rozdzielone rzekami, które dla wielu amazońskich ptaków stanowią barierę geograficzną, bo ptaki po prostu nie lubią latać nad wodą. Tutaj jest ich geograficzna dystrybucja z artykułu:
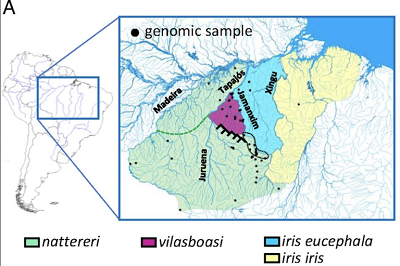
Można zobaczyć, że LV, przypuszczalny “hybrydowy gatunek”, leży pomiędzy dwoma rodzicielskimi populacjami LN i LI (dwoma podgatunkami). Kiedy po raz pierwszy znaleziono LV w 1957 r., sądzono, że jest to tylko “strefa hybrydowa”: obszar hybrydyzacji między dwoma gatunkami rodzicielskimi.
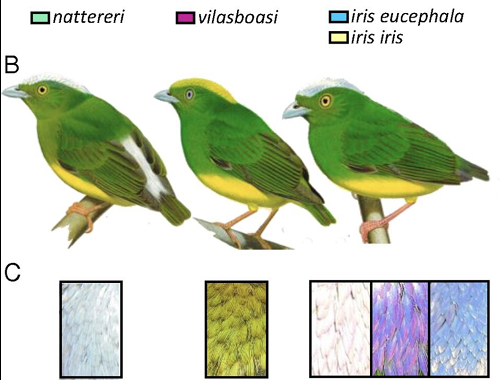
Te trzy gatunki różnią się najbardziej wyraźnie kolorem korony na czubku głowy. Jak piszą w artykule:
Czubek głowy L. iris jest opalizujący i waha się od lśniąco białego (jego typowy wygląd, który jest bardzo podobny do L. nattereri) do niebieskiego lub fioletowego, zależnie od kąta padania światła. Samce dwóch podgatunków L. iris rozmieszczonych po obu stronach rzeki Xingu mają niemal identyczne upierzenie, z L. iris iris mającym cienki zielony pas między górną szczęką a koroną i z koroną rozciągającą się aż do szczęki L. iris eucephala. Samice (nie pokazane) wyglądają jak samce, ale brak im kontrastu między ubarwieniem korony i kupra, i nie różnią się specjalnie między gatunkami.
W odróżnieniu od tego, LV ma żółtą koronę, bardzo różniącą się od obu gatunków rodzicielskich (patrz poniżej).
Tutaj jest LV (“gorzyczek zielonkawy”) ze swoją żółtą koroną:

I LN (“gorzyczek białorzytny”) z białą koroną:

No cóż, jakie są dowody, że LV powstał z hybrydyzacji między LN i LI? Reszta jest genetyką. Genom LV jest mieszanką genów dwóch rodzicielskich gatunków i próba rekonstrukcji przodków tych ptaków przy pomocy zarówno standardowej filogenetyki, jak “koalescencyjnej” symulacji, pokazuje, że jest dużo bardziej prawdopodobne, że LV powstał z mieszania genów LN i LI niż, że oddzielił się od jednego lub drugiego gatunku (konwencjonalny scenariusz “odgałęziania się” w specjacji). Między 15% a 38% genomu LV pochodzi od LN a 62%-85% genomu od LI, zależnie od metody użytej do obliczeń.
Jednym problemem jest to, że LV może nadal reprezentować izolowaną populację hybrydową, czyli strefę hybrydową, między dwoma gatunkami. Autorzy mówią, że nie jest to prawdopodobne, ponieważ czyste hybrydy F1 tych dwóch gatunków (tj. pierwszego pokolenia) miałyby 50% genów każdego gatunku zamiast tego skrzywionego rozkładu. Tak jednak byłoby tylko, gdyby hybrydy F1 były bezpłodne i nie mogłyby tworzyć dalszych hybryd przez rozmnażanie się. Jest nadal możliwe, że mamy tutaj strefę hybrydową, której członkowie nie są izolowani reprodukcyjnie od dwóch gatunków rodzicielskich, a więc LV nie jest naprawdę “gatunkiem”. Kluczowym dowodem jest, czy LV są izolowane reprodukcyjnie od LN i LI (już są izolowane geograficznie, ale to nie liczy się jako bariera reprodukcyjna w “pojęciu biologicznego gatunku”, bo w takim przypadku każda izolowana geograficznie populacja byłaby gatunkiem).
Czy więc LV są izolowane reprodukcyjnie od obojga rodziców? Nie wiemy tego z pewnością, bo nie zamieszkują tego samego terenu. Barrera-Guzmán i in. podejrzewają, że LV byłyby izolowane – przez inaczej ubarwione głowy. Ptaki używają koloru korony na głowie do identyfikowania partnerów własnego gatunku i do odrzucania niewłaściwych, więc autorzy sądzą, że samce LV nie zostałyby uznane za odpowiednich partnerów przez samice LN i LI z powodu ich żółtych głów. Podobnie, jest możliwe, że samice LV nie rozpoznałyby opalizujących głów gatunków rodzicielskich, więc izolacja reprodukcyjna działałaby na wszystkich frontach.
Ciekawą rzeczą z koroną LV jest jej nowy, żółty kolor. Autorzy odkryli, że opalizacja koron LN i LI powodowana jest przez różne właściwości strukturalne ich piór i że korony LV są czymś pośrednim między tymi dwoma zestawami właściwości. Ciekawe, że jeśli tworzysz czystą hybrydę F1 między LN i LI, otrzymujesz burą koronę o tych samych pośrednich właściwościach strukturalnych, jak hybryda LV, ale nie jest ona żółta!
Autorzy następnie tworzą – rozsądnie, moim zdaniem – następujący scenariusz: dwa gatunki rodzicielskie stworzyły hybrydy, dając populację, która mieszkała w lesie (gdzie żyje LV), ale z burymi koronami. Dobór płciowy lub dobór naturalny spowodował następnie ewolucję struktur zatrzymujących karetonoidy w populacji hybryd, żeby samice widziały samce w lesie. (Preferencje samic do żółtego koloru podążałyby – przez dobór płciowy – razem z tym.) Tak więc sama hybrydyzacja nie wystarczyła do specjacji: izolacja reprodukcyjna musiałaby obejmować jakiś dobór po hybrydyzacji.
Wnioski: Sądzę, że autorzy mają dobry argument, że odkryli rzadką specjację u ptaków przez diploidalną hybrydyzację. Nie jest absolutnie przekonujące, ponieważ nadal jest możliwe, że mamy tu chmarę hybryd, która nie jest nowym gatunkiem i nie jest izolowana reprodukcyjnie od rodziców. Jeśli potrafią pokazać silną dyskryminację w doborze partnerów na bazie koloru korony (lub czegokolwiek innego), dane będą bardziej przekonujące. Nadal jednak sądzę, że odkryli możliwy (wręcz prawdopodobny) przypadek, który z niejaką dalszą pracą może reprezentować jeden z nielicznych przypadków gatunku diploidalnych hybryd u zwierząt.
Niektórzy ludzie mogą powiedzieć: “no cóż, to jest nowy ewolucyjny paradygmat, bo specjacja ma zdarzać się przez rozgałęzianie linii rodowych, a nie przez anastomozę (połączenie) odrębnych gałęzi w jedną”. Zgadza się, gdyby to był sposób, w jaki normalnie powstają gatunki, uzasadniałoby to dość poważną rewizję darwinowskiej teorii. Dowody wskazują jednak na to, że nie jest to sposób powstania większości diploidalnych gatunków. Gdyby tak było filogenetyczna analiza nie rozwiązałaby niczego z drzewa życia: mielibyśmy tylko przypominającą spaghetti masę. U Drosophila, grupy, którą znam najlepiej, tak najwyraźniej nie jest: regułą jest odgałęzianie, nie zaś łączenie. Chociaż jest trochę więcej przepływu genów między gatunkami zwierząt niż podejrzewaliśmy dwadzieścia lat temu, jest to „introgresja”, nie specjacja. Nadal nie widzimy dowodów, że tworzenie się nowych gatunków zwierząt zachodzi przez łączenie się już istniejących gałęzi (gatunków).
______
Barrera-Guzmán, A. O., A. Aleixo, M. D. Shawkey, and J. T. Weir. 2018. Hybrid speciation leads to novel male secondary sexual ornamentation of an Amazonian bird. Proc. Nat. Acad. Sci. USA 115:E218-225. Published ahead of print December 26, 2017, doi:10.1073/pnas.1717319115
Hybrid speciation in Amazonian manakins
Why Evolution Is True, 14 stycznia 2018
Tłumaczenie: Małgorzata Koraszewska
Profesor (emeritus) na wydziale ekologii i ewolucji University of Chicago, jego książka "Why Evolution is True" (Polskie wydanie: "Ewolucja jest faktem", Prószyński i Ska, 2009r.) została przełożona na kilkanaście języków, a przez Richarda Dawkinsa jest oceniana jako najlepsza książka o ewolucji. Jerry Coyne jest jednym z najlepszych na świecie specjalistów od specjacji, rozdzielania się gatunków. Jest wielkim miłośnikiem kotów i osobistym przyjacielem redaktor naczelnej.