Dlaczego ryby jaskiniowe ewoluują tak, by oślepnąć?
Najpierw przykład: Smith pokazuje klasyczny przypadek Astyanax mexicanus, lustrzenia meksykańskiego lub “ślepczyka jaskiniowego” znajdowanego w jaskiniach na południowym zachodzie USA i w północnym Meksyku. Ślepczyk jaskiniowy jest uważany za ten sam gatunek jak jego żyjąca na powierzchni forma. Poniżej są zdjęcia każdej z jego form oraz zasięg ślepych ryb (z artykułu). Istnieje 26 znanych populacji ślepej formy, reprezentujące co najmniej pięć niezależnych wypadków ewolucyjnej utraty oczu. Eksperymenty pokazały, że odmiany jaskiniowa i powierzchniowa są wzajem ze sobą płodne i że utrata oczu u postaci jaskiniowej dotyczy kilku genów, a nie tylko jednego:

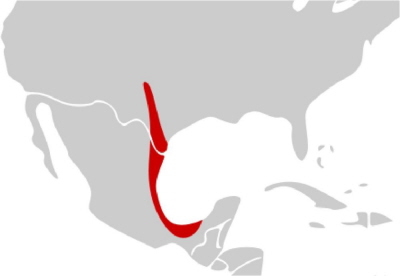
Forma jaskiniowa odżywia się głównie kożuchem bakteryjnym na wodzie, który tworzy się przy rozkładzie odchodów nietoperzy i świerszczy. Oczy istnieją nadal jako szczątkowe pozostałości pod powierzchnią skóry, ale zaczynają rozwój jako normalne oczy, a potem cofają się w miarę dorastania ryby. (Już to jest dowodem na ewolucję.) Ryby z przynajmniej jednej jaskini mają zdolność wykrywania światła, ale inne nie mają takiej zdolności; prawdopodobnie odzwierciedla to różne etapy ewolucyjne utraty oczu (lub też inne poziomy światła w jaskiniach). Ryby orientują się w przestrzeni dzięki wibracjom. Jak pisze Smith: „Naukowcy łapali te ryby po prostu przez zanurzanie sieci w wodzie i wprawianiu jej w wibracje”.
Tutaj są trzy hipotezy Smitha w sprawie ewolucji utraty oczu. Jego słowa są wcięte (podkreślenia są moje). Twierdzę, że dwie z tych hipotez są połączone, jedna jest w dużej mierze niepoprawna, a także pomija on jeszcze inną hipotezę.
Według pierwszej hipotezy utrata oczu istotnie jest spowodowana przez bezpośredni dobór naturalny ponieważ istnieją korzyści nie posiadania oczu w ciemności. Badania pokazały, że utrzymanie tkanki ocznej, szczególnie siatkówki i związanej z tym tkanki nerwowej pociąga za sobą wysoki koszt metaboliczny. (Moran et al., 2015; Protas et al., 2007). Dlatego ryby jaskiniowe bez oczu mają przewagę w tym środowisku, gdzie źródła energii (pokarm) są rzadkie, ponieważ ślepa ryba nie marnuje energii na niepotrzebne narządy.
Jest to rozsądna hipoteza i tę hipotezę zawsze najpierw podawali moi studenci, kiedy ich o to pytałem. Stosuje się ona do zanikania wszystkich nieużywanych narządów, jak maleńkie wzgórki, które są szczątkowymi „skrzydłami” kiwi. „Niemarnowanie energii” sugeruje oczywiście, że energia będzie kierowana na inne narządy lub funkcje, które podnoszą reprodukcję, bo to kryje się w powiedzeniu, że zredukowanie oczu daje rybie jaskiniowej przewagę drogą doboru naturalnego.
Druga hipoteza posługuje się zjawiskiem plejotropii, to jest, sytuacjami, w których mutacja w jednym genie wpływa na więcej niż jedną cechę fenotypową. Na przykład, istnieją dowody, że jeden z genów odpowiedzialnych za utratę oczu ryb jaskiniowych podnosi także liczbę kubków smakowych, co pomaga rybie jaskiniowej w skuteczniejszym znajdowaniu pokarmu (Gross, 2012). Dobór naturalny na wzrost liczby kubków smakowych promowałby więc także ślepotę.
Moim zdaniem druga hipoteza nie różni się zasadniczo od pierwszej. Przecież, jeśli zasoby są skierowane od unieczynnionych genów do innych narządów lub funkcji, które podnoszą reprodukcję, to te inne cechy będą odbijać efekty plejotropowe mutacji, która zredukowała oczy. Nie widzę materialnej różnicy między a) redukujący oczy gen podnosi liczbę kubków smakowych (hipoteza „plejotropowa”); lub b) redukująca oczy mutacja powoduje, że więcej składników odżywczych jest dostępnych dla innych narządów przez zredukowania wymogów energetycznych na budowę oczu. W obu wypadkach mutacja redukująca tworzenie się oczu ma korzystne skutki dla innych aspektów rozwojowych. Jedno i drugie to przypadki „plejotropii”.
Trzecia hipoteza oparta jest na mutacji neutralnej i dryfie genetycznym. Aż nazbyt często podręczniki używają wymiennie terminów „ewolucja” i „dobór naturalny”, ignorując znaczenie dryfu genetycznego. Dryf genetyczny jest „procesem zmiany w składzie genetycznym populacji z powodu przypadku lub wydarzeń losowych zamiast z powodu doboru naturalnego, z czego wynikają fluktuacje częstości występowania danego allelu” (Biology Online, 2008). Dryf genetyczny różni się od doboru naturalnego, ponieważ obserwowane zmiany w częstości występowania danego allelu są całkowicie przypadkowe, nie są zaś wynikiem doboru naturalnego dla jakiejś cechy. Dryf genetyczny może mieć stosunkowo duży wpływ w małych populacjach, takich jak typowa populacja ryb jaskiniowych. Według hipotezy mutacji neutralnej i dryfu genetycznego normalny proces mutacji w małej populacji ryb jaskiniowych czasami daje mutacje neutralne (mutacje, które prowadzą do zmian fenotypowych, na które nie działa dobór naturalny) i pod nieobecność doboru naturalnego całkowicie przypadkowe wydarzenia mogą czasami spowodować wzrost częstości występowania takiej mutacji. Takie zmiany mogą obejmować degenerację oczu.
To rozumowanie nie jest poprawne. Nawet gdyby geny redukujące oczy były neutralne i nie dawały rybom bez oczu przewagi reprodukcyjnej, dryf genetyczny (losowe fluktuacje form z oczyma i bez) same nie mogłyby spowodować powszechnej utraty oczu w jaskiniach, a jaskinie zawierają tylko ryby bez oczu. Dryf genetyczny daje „losowy” efekt: zmienną mieszankę ryb z oczyma i bez w różnych jaskiniach. A tego nie widzimy.
Dryf genetyczny może odgrywać niewielką rolę w utracie oczu (mutacje tylko trochę szkodliwe mają większą szansę przetrwania w małych populacjach), ale sądzę, że Smith pomija tutaj zjawisko nielosowe: kierunkową mutację. Nie mam na myśli tego, że w jakiś sposób zwiększa się częstość występowania w jaskiniach mutacji, które wyłączają oczy w porównaniu do populacji na powierzchni – to byłby proces lamarckowski albo teleologiczny – ale losowe mutacje dzieją się zarówno w populacjach w jaskini, jak na powierzchni. W populacjach na powierzchni mutacje, które redukują lub wyłączają oczy są wyplenione przez dobór naturalny, a takie mutacje są liczniejsze niż mutacje tworzące lepsze oczy. Pamiętajmy, że w genach na tworzenie się oczu, jak i we wszystkich genach, losowe uderzenie w skomplikowaną i wyewoluowaną sekwencję DNA ma większe prawdopodobieństwo uszkodzenia genu niż poprawienia jego wpływu na reprodukcję.
Dlatego, z potokiem mutacji wpływających na oczy obu populacji i ogólnie degenerujących oczy, liczniejsze “złe” mutacje będą eliminowane w populacji na powierzchni, ale nie będzie doboru przeciwko nim w populacji w jaskini, a więc będą się akumulować – być może wspierane przez dobór naturalny (hipoteza 1 i 2 powyżej). Spójrz na to w ten sposób: jeśli masz park samochodów, którymi nigdy nikt nie jeździ, ale ludzie losowo regulują silniki tych samochodów, niczego o nich nie wiedząc, wszystkie nieużywane samochody w końcu stracą zdolność jazdy. Jest tak, ponieważ losowe regulowanie silnika z większym prawdopodobieństwem uszkodzi go niż poprawi jego funkcjonowanie. Silnikami są oczy ryb jaskiniowych, a mutacje są regulowaniem. Regulowanie akumuluje się, ponieważ samochody nie muszą jeździć. Sądzę, że jest to bardziej prawdopodobne wyjaśnienie niż prosty dryf genetyczny, który i tak wydaje się nieprawdopodobny, ponieważ mutacje redukujące oczy przypuszczalnie nie są „neutralne” z powodów podanych powyżej, ale także z powodu tego, co mam do powiedzenia w mojej czwartej hipotezie poniżej.
Smith mówi, że:
. . . badania sekwencji innych genów związanych z oczyma ryb jaskiniowych pokazują wysoką częstość substytucji zarówno w odcinkach kodujących, jak niekodujących, co popierałoby hipotezę dryfu genetycznego (Retaux & Casane, 2013).
Wydaje się to jednak być błędne z kilku powodów. Po pierwsze wysoka częstość występowania substytucji w odcinkach kodujących może być spowodowana każdym rodzajem doboru naturalnego omawianym powyżej. Po drugie, odcinki niekodujące (części DNA, które nie kodują białek) mogą czasami wpływać na ekspresję i regulację genów. Co ważniejsze, nie mogłem znaleźć żadnych danych w pracy Retaux i Casne’a sugerujących wyższą częstość występowania prawdziwie neutralnych, niekodujących mutacji u tych ryb jaskiniowych. (Mogłem to przeoczyć, ale nie wydaje się, by to tam było.) Tutaj jest ten akapit (uwaga: to jest dla genetyków ewolucyjnych):
Raporty cytowane powyżej dotyczą tylko ewolucji sekwencji kodujących. Jednak ewolucja fenotypowa (włącznie z utratą struktur) może także zachodzić przez zmiany w sekwencjach niekodujących, cis-regulatorowych. Wśród słynnych przykładów jest utrata kości miednicy u cierników słodkowodnych przez delecję wzmacniacza transkrypcji Pitx1 [98, 99], lub zyskanie albo utrata wzorów pigmentacji u Drosophilae przez kooptowanie lub mutacje elementów regulatorowych w genie pigmentacji yellow [100]. Chociaż dokładny mechanizm nie jest znany, to zdarzyło się w krystalinie αA u jaskiniowego Astyanax [55, 101]. Ta opiekuńcza i przeciwapopotyczna krystalina, której kodująca sekwencja jest niemal identyczna u ryb powierzchniowych i jaskiniowych (różnica tylko jednego aminokwasu), ma silnie wyhamowaną ekspresję w soczewkach ryb jaskiniowych podczas rozwoju i sugeruje się to jako potencjalnego głównego gracza w początkach apoptozy soczewek u ryb jaskiniowych. U golców Heterocephalus glaber, gamma-krystaliny są wyłączone po urodzeniu [46]. U golca Spalax ehrenbergi, promotor αB-krystaliny i odcinki między genami selektywnie tracą aktywność w soczewkach po 13,5 dniach [102, 103]. Te przykłady pokazują jak zmiany w sekwencjach regulatorowych zachodzą także w zwierzętach jaskiniowych i innych zwierzętach podziemnych.
Proszę zauważyć, że nie ma tutaj danych o “wysokiej częstości substytucji […] w odcinkach niekodujących” oczu ryb jaskiniowych. Widzimy zmiany w regulacji genów bez towarzyszących im zmian w sekwencji regulowanych genów, ale to jest prawdopodobnie spowodowane „kodującymi” zmianami w innych genach regulatorowych lub substytucjami w odcinkach regulatorowych, które nie są „neutralne”, ponieważ wpływają na tworzenie się oczu. (Proszę zauważyć, że Smith podkreśla „neutralne” mutacje w swojej trzeciej hipotezie.) Te odcinki regulatorowe są więc przedmiotem doboru naturalnego i nie są zmianami „neutralnymi”, na które działa wyłącznie dryf genetyczny, nawet jeśli są niekodujące. Istotnie, oczekiwalibyśmy, że dobór dostarczy tej obserwacji: więcej substytucji gromadzących się w odcinkach regulatorowych w rybie jaskiniowej niż powierzchniowej! Nie ma tutaj potrzeby dryfu.
Czwarta hipoteza Coyne’a (właściwie nie moja, ale pominięta przez Smitha). Oczy są delikatnymi narządami, łatwo je uszkodzić i mają skłonność do infekcji. Jeśli zredukujesz oczy, kiedy ich nie potrzebujesz, masz mniejszą skłonność do odnoszenia takich uszkodzeń środowiskowych, a więc geny redukujące oczy czynią, że ich nosiciele mają większe szanse na przeżycie i rozmnożenie się. Tak, to jest rodzaj utraty oczu promowany przez dobór, ale jest pojęciowo inny od hipotezy 1 i 2 powyżej. Chciałbym, by Smith wspomniał o tym także.
Na koniec Smith mówi, że wszystkie te sugerowane procesy mogły działać razem:
A więc, która odpowiedź jest poprawna? Jakie są dowody genetyczne na poparcie każdej z tych hipotez? Jak często dzieje się w nauce, odpowiedzią jest prawdopodobnie, że te wyjaśnienia nie wykluczają się wzajemnie; jest prawdopodobne, że wszystkie trzy częściowo wyjaśniają ślepotę ryb jaskiniowych. Dla zrozumienia tego musimy mieć trochę dalszych informacji o genetyce A. mexicanus.
No cóż, wyjaśnienia mogą nie wykluczać się wzajemnie, ale nie ma uzasadnienia do twierdzenia, że wszystkie trzy „prawdopodobnie” wyjaśniają ślepotę ryb jaskiniowych. Jedna lub dwie hipotezy mogą wyjaśnić większość problemu utraty oczu. Sam fakt, że istnieje kilka możliwości, nie oznacza, że działały zgodnie.
Choć próbuję poprawić lub dorzucić swoje trzy grosze do pracy Smitha, nie próbuję powiedzieć, że jest to zły artykuł. Nie jest: porusza ważny temat do omawiania na lekcjach ewolucji i sugeruje bogactwo hipotez i eksperymentów. Zawiera też bardzo użyteczne sugestie, jakie błędne pojęcia mogą mieć na ten temat studenci i jak je poprawnie wyjaśnić. Po prostu uważam, że myśli powinny być formułowane i wyrażane staranniej. Choć nie znamy dokładnych przyczyn ewolucyjnych utraty oczu u lustrzeni meksykańskich, fakt, że zdarzyło się to niezależnie kilka razy, jak również zdarzyło się u innych gatunków żyjących w jaskiniach, sugeruje, że główną rolę odgrywał tu raczej dobór naturalny niż dryf genetyczny.
___________
Smith, M. U. 2017. How does evolution explain blindness in cavefish? The American Biology Teacher, Vol. 79 No. 2, February 2017; (pp. 95-101) DOI: 10.1525/abt.2017.79.2.95
Why do cave fish evolve to become blind
Why Evolution Is True, 23 kwietnia 2017
Tłumaczenie: Małgorzata Koraszewska

Profesor (emeritus) na wydziale ekologii i ewolucji University of Chicago, jego książka "Why Evolution is True" (Polskie wydanie: "Ewolucja jest faktem", Prószyński i Ska, 2009r.) została przełożona na kilkanaście języków, a przez Richarda Dawkinsa jest oceniana jako najlepsza książka o ewolucji. Jerry Coyne jest jednym z najlepszych na świecie specjalistów od specjacji, rozdzielania się gatunków. Jest wielkim miłośnikiem kotów i osobistym przyjacielem redaktor naczelnej.