Nareszcie odkryta mutacja krępaka nabrzozaka

Dlaczego zmiana ewolucyjna? (Pamiętajmy, ewolucję można zdefiniować jako “zmianę genetyczną w populacji”). Hasło w Wikipedii “Peppered moth evolution” daje dobre streszczenie.
Wczesne eksperymenty pokazały, że prawdopodobną przyczyną było żerowanie ptaków. Ptaki są drapieżnikami kierującymi się wzrokiem i zbierają ćmy z drzew. Przed rewolucją przemysłową drzewa brytyjskie miały jasny kolor, często ozdobione były porostami, a więc jasne ćmy były zakamuflowane i mniej narażone na zjedzenie niż ciemne. Pierwszą ciemną ćmę opisano jako rzadkość w 1848 r. w Manchesterze, a potem częstość występowania ciemnej odmiany rosła raptownie, kiedy sadze z kominów fabrycznych przyciemniły drzewa i zabiły porosty.
Ta zmiana koloru drzew, najbardziej wyraźna na terenach przemysłowych, takich jak Manchester, odwróciła przewagę selekcyjną odmian ciemnej i jasnej: ciemne ćmy były teraz lepiej zakamuflowane i mniej zagrożone zjedzeniem. Częstość występowania tej odmiany bardzo wzrosła w wielu miejscach: do niemal 100% w ponurym Manchesterze (przepraszam, Matthew). Wszystkie te zmiany zaszły w ciągu około 50 lat, a więc dobór naturalny był bardzo silny.
Tutaj jest zdjęcie pokazujące odmiany jasną i ciemną na drzewach przyciemnionych i “normalnych”. Można zobaczyć, jak ćmy niewłaściwego koloru wyróżniają się i byłyby oczywiście widoczne dla ptaków szukających posiłku.
Przyciemnione sadzą drzewo:

Drzewo nie narażone na zanieczyszczenie. Proszę zauważyć nakrapiane barwy odmiany typica, które zlewają się z korą drzewa:

Eksperymenty Bernarda Kettlewella w latach 1950., w których umieszczał ćmy na drzewach a potem ponownie je chwytał, sugerowały hipotezę ptaków drapieżnych, ponieważ rzadziej łapał ponownie ciemne ćmy umieszczone w jasnych lasach niż jasne ćmy, co sugerowało, że niewłaściwie ubarwione ćmy częściej ginęły. Było odwrotnie, kiedy umieszczał ćmy w zanieczyszczonych lasach. Inne badania, w których także umieszczano ćmy na drzewach, sugerowały ten sam scenariusz.
Były jednak problemy z zaprojektowaniem i wykonaniem tych eksperymentów i w 1998 r. narobiłem odrobinę szumu przez wskazanie tych problemów oraz powiedzenie, że pokazanie, iż przyczyną zmiany są ptaki drapieżne, jest sugestywne, ale nie rozstrzygające. Brytyjscy ewolucjoniści, niezmiernie przywiązani do tej sprawy, nieco mnie demonizowali, a kreacjoniści, oczywiście, podchwycili moje zastrzeżenia i nadużyli ich do rzucania wątpliwości na całą ewolucję. Jeśli podręcznikowy przykład doboru naturalnego jest wadliwy, twierdzili, to całe przedsięwzięcie darwinowskie jest błędne.
Jednak Michael Majerus, badacz z Cambridge, od 2001 r. zaczął powtarzać porządnie eksperymenty uwolnienia i ponownego złapania (używał tylko jasnych lasów, bo ciemnych, zanieczyszczone już nie było) i otrzymał te same wyniki, jakie otrzymał Kettewell: w tych jasnych lasach ciemne ćmy, wypuszczone razem z jasnymi, były mniej często chwytane ponownie i Majerus widział ptaki zjadające te ćmy. (Niestety, zmarł zanim praca została opublikowana, ale jego koledzy napisali artykuł i opublikowali go.) Wyniki Majerusa wraz z obserwacją, że ten sam gatunek w Ameryce Północnej doświadcza takich samych zmian w częstości występowania odmian w miarę wzrostu i spadku zanieczyszczeń w USA, przekonały mnie, że historia ciem jest zupełnie solidna. Nie ma wątpliwości, że mamy tu przypadek ewolucji drogą doboru naturalnego w działaniu i wiemy, jaki rodzaj selekcji zachodzi.
Wiedzieliśmy od dziesięcioleci, że różnica między odmianami carbonaria i typica jest spowodowana zmianami w jednym genie, bo krzyżowanie pokazywało segregację mendlowską, z carbonaria zachowującym się jako allel dominujący. Nie wiedzieliśmy jednak, o który gen chodzi, chociaż w ostatnich latach ograniczono to do liczącego 400 tysięcy zasad odcinka genomu ćmy. Chociaż fakt, że jest to przypadek ewolucji drogą doboru naturalnego, nie zależy od wiedzy, który dokładnie gen bierze w tym udział, dla pełnej historii od genu do koloru do sił ekologicznych (drapieżnictwo ptaków) miło byłoby znać ten gen.
Duża grupa badaczy donosi obecnie w “Nature”, że znaleźli ten gen. W artykule autorstwa Arjen E. van’tHof i in. (odnośnik poniżej) identyfikują gen powodujący różnice barw ciem jako cortex, dobrze znany gen, który badano u muszek owocowych (Drosophila). Towarzyszący mu artykuł w tym samym numerze „Nature” autorstwa Nicola Nadeau i in. (odnośnik poniżej) pokazuje, że mutacje w cortex biorą udział w tworzeniu wzorów i w mimikrze u wielu gatunków Lepidoptera.
Tutaj są istotne rezultaty pracy van’t Hoff i in.:
- Gen, w którym mieszczą się zmiany powodujące, że jasne ćmy stają się ciemne, nazywa się cortex, i wiadomo, że u Drosophila bierze udział w podziale komórek jajeczek samic. Nie znano, przynajmniej u much, jego roli w zmianie ubarwienia.
- Związek między cortex a ubarwieniem odkryto dzięki “mapowaniu asocjacyjnemu”. Badacze wzięli grupy ciem obu odmian i sekwencjonowali DNA w regionie, gdzie znajdowała się mutacja, szukając systematycznej zmiany w DNA, która odróżniałaby w sposób niemal doskonały te odmiany.
- Znaleziona zmiana nie była mutacją jednej zasady w DNA, ale wstawieniem “transpozonu”, sekwencji DNA, która potrafi poruszać się w genomie, do odmiany carbonaria. Cała wstawiona sekwencja składa się z 21 925 nukleotydów i wskoczyła do genu cortex w całości.
- Transpozon uaktywnił (zamiast wyciszyć) gen cortex, podnosząc ilość jego produktu w sposób, którego nie rozumiemy. Co dziwne, wzmożona aktywność genu w ciemnej odmianie jest znacznie silniejsza u larw (gąsienic) niż u poczwarek i dorosłych, przypuszczalnie dlatego, że początki łusek i ich kolor formują się na tym stadium rozwojowym.
- Związek między transpozonem a barwą był niemal doskonały, ale nie stuprocentowy. Wszystkim odmianom typica i insularia brakowało tego element, podczas gdy 105 z 110 czarnych carbonaria miało te transpozony. Znaczy to, że inne geny poza cortex mogą wpływać na kolor – lub mogą być efekty rozwojowe/środowiskowe, które nie są genetyczne – ale że wstawienie transpozonu do cortex jest prawdopodobnie najważniejsze – ono produkuje paliwo dla doboru naturalnego ubarwienia.
- Przyglądając się DNA wokół transpozonu badacze mogli ocenić wiek tej mutacji. Gdyby powstała niedawno u jednego osobnika, większość nosicieli tej mutacji miałaby podobne sekwencje w pobliżu, bo rekombinacja nie miałaby czasu na zestawienie transpozonu z pobliskim DNA innych osobników. Gdyby mutacja była stara, otaczające ją odcinki byłyby bardziej zróżnicowane, bo rekombinacja połączyłaby wstawiony gen z pobliskimi genami różnych osobników. Używając symulacji badacze podali rok 1819 jako najbardziej prawdopodobną datę mutacji, czyli na krótko zanim zobaczono ją po raz pierwszy w naturze. „Rozstęp ćwiartkowy” dla dat, jak rozumiem, oznacza prawdopodobieństwo między 25% a 75% powstania tego w latach 1681-1806. Poniżej jest wykres pokazujący gęstość prawdopodobieństwa, kiedy zaszła mutacja carbonaria (tj., kiedy przeskoczył transpozon). Można zobaczyć, że wszystkie te daty są stosunkowo niedawne. Najwyższe prawdopodobieństwo jest przy roku 1819, niedaleko od pierwszego zobaczenia ciemnej ćmy (1848 r.; linia kropek). Brytyjczycy, zapaleni kolekcjonerzy motyli, z pewnością szybko znaleźliby nową odmianę!
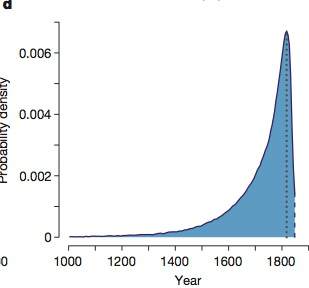
Wynik. Jak powiedziałem, uważam tę pracę za nagrodę na deser: znaliśmy już główne punkty historii – że podstawą zmiany adaptacyjnej w ubarwieniu motyla w oparciu o drapieżnictwo ptaków był jeden gen. Teraz wiemy, że tym genem jest cortex.
Wiemy także, że zmianę w cortex, powodującą zmianę ubarwienia ćmy z jasnej na ciemną, spowodowało wstawienie ruchomego transpozonu, nie zaś „konwencjonalna” zmiana w jednej zasadzie sekwencji DNA. Jest to nadal „mutacja”, ale duża, zmieniająca ilość produktu genu. Mogą istnieć inne adaptacje oparte o wstawianie lub usuwanie transpozonów, ale ja ich nie znam. (Czytelnicy, którzy posiadają taką wiedzę, powinni o tym napisać poniżej.)
Mutacja pojawiła się na krótko przed zmianą środowiska – zanieczyszczeniem – która spowodowała ewolucję różnic ubarwienia. Jest to interesujące, ale nie było konieczne, bo takie mutacje pojawiają się nieustannie i mogą pozostawać w populacji na zawsze. Choć bowiem dobór naturalny wyplenia takie geny z populacji (ciemne ćmy byłyby upośledzone przed rewolucją przemysłową) mutacja wstawia je z powrotem, a więc rezerwuar rzadko występujących mutacji, które istnieją w populacji, może być podstawą nowych adaptacji, jeśli zmieni się środowisko. (Nazywa się to „równowagą mutacji/selekcji”. Pamiętajmy jednak, że te mutacje nie pozostają w populacji w celu dostarczenia przyszłej ewolucji adaptacyjnej. Błędy w DNA zdarzają się losowo, nie mogą przewidywać przyszłych potrzeb organizmu i czasami, choć rzadko, okazują się użyteczne.)
Na koniec: ten artykuł oraz artykuł o Heliconius pokazują, że choć muszki owocowe są dobrym „organizmem modelowym” – gatunkiem, który wiele nauczył nas o rozwoju i genetyce (większość tego co wiemy o genetyce mendlowskiej, zostało wypracowane na Drosophila) – nie mówią nam wiele o ewolucji Biston betularia. Jest tak z prostego powodu: muchy nie mają łusek, a więc nie można używać Drosophila do badania koloru łusek. W tym wypadku, co widzimy tak często w ewolucji, gen, który robi coś w jednym gatunku, może zostać dokooptowany do całkiem innej funkcji w drugim gatunku. Mogliśmy to zrozumieć dopiero, kiedy zdobyliśmy nowe narzędzia genetyczne i rozwojowe w ciągu ostatnich 30 lat.
h/t: Matthew Cobb, Jonathan
_________
van’t Hof, A. et al. 2016. The industrial melanism mutation in British peppered moths is a transposable element. Nature 534:102-105.
Nadeau, N. J. et al. 2016. The gene cortex controls mimicry and crypsis in butterflies and moths. Nature 534:106-110.
Peppered moth mutation discovered at last
Why Evolution Is True, 3 czerwca 2016
Tłumaczenie: Małgorzata Koraszewska

Profesor na wydziale ekologii i ewolucji University of Chicago, jego książka "Why Evolution is True" (Polskie wydanie: "Ewolucja jest faktem", Prószyński i Ska, 2009r.) została przełożona na kilkanaście języków, a przez Richarda Dawkinsa jest oceniana jako najlepsza książka o ewolucji. Jerry Coyne jest jednym z najlepszych na świecie specjalistów od specjacji, rozdzielania się gatunków. Jest wielkim miłośnikiem kotów i osobistym przyjacielem redaktor naczelnej.