Dlaczego seks? Eksperymenty na muszkach owocowych sugerują, że wyewoluował, by opierać się infekcji

Można pokazać, że w rzeczywistości przy typowym dla seksu procesie: rekombinacji (chromosomach wymieniających odcinki z drugim chromosomem z pary) i segregacji (różne odcinki par chromosomów zestawiają się w jajeczku lub plemniku) jest znaczna strata ewolucyjna. Strata jest podwójna (nazywa się to „kosztem seksu” lub „kosztem posiadania samców”), a więc osobnik, który potrafi rozmnażać się bezpłciowo, pozostawia dwukrotnie więcej kopii swoich genów niż osobnik, który uprawia seks. Innymi słowy, gen na eliminację płci i rozmnażanie bezpłciowe powinien rozejść się w populacji jak burza. Według teorii ewolucyjnej każdy gatunek powinien rozmnażać się bezpłciowo!
Tak jednak nie jest. Olbrzymia większość nie-mikrobowych gatunków na Ziemi rozmnaża się płciowo. Biorąc pod uwagę koszty, musi być w tym jakaś olbrzymia korzyść ewolucyjna, która jest wystarczająco silna, by przeważyć dwukrotny koszt seksu.
Niestety, ewolucjoniści nie wiedzą, co to jest za korzyść, więc ewolucyjne wyjaśnienie wszechobecności seksu nadal jest zagadką. Jest to jedna z wielkich Czarnych Skrzynek w mojej dziedzinie.
Jednym z wyjaśnień, które ma pewne poparcie, jest, że seks umożliwia tworzenie genetycznie bardziej zróżnicowanego potomstwa. Zróżnicowanie to jest rezultatem rekombinacji i segregacji w genomach rodziców. Jeśli ma się bardziej zróżnicowane potomstwo, być może część potomków będzie miała właściwą kombinację genetyczną, by oprzeć się infekcjom i innym wyzwaniom środowiskowym. Jeśli jednak całe potomstwo jest dokładnie takie samo jak rodzic, jak to jest w większości przypadków rozmnażania bezpłciowego, to nie ma żadnej szansy na różnorodność. Jeśli rodzic nie może oprzeć się infekcji, to nie może jej oprzeć się także jego potomstwo. To może być naciskiem selekcyjnym na geny, które dają rozmnażanie płciowe, pozwalając na przeżycie części potomstwa.
Jest to tak zwana “hipoteza Czerwonej Królowej” na rzecz ewolucji seksu. Nazwa zaczerpnięta jest ze sceny w Po drugiej stronie lustra, w której Alicja i Czerwona Królowa biegną, ale zawsze pozostają w tym samym miejscu. Nazwa jest właściwa, bo jeśli hipoteza jest poprawna, organizmy ciągle biegną, by wyprzedzić infekcje pasożytnicze; niezależnie jednak od tego, jak bardzo zróżnicowane jest potomstwo, zawsze znajdzie się jakiś nowy pasożyt (albo nowa mutacja pasożyta), a więc nacisk na zachowanie płciowości pozostaje.
Można pokazać, że w pewnych warunkach ten wzrost różnorodności wśród potomstwa może selekcjonować na rzecz rozmnażania płciowego – to jest, korzyści z rekombinacji i segregacji oraz łączenia jajeczka lub plemnika z gametą innego osobnika mogą przeważyć dwukrotny koszt seksu. Dowodem na to, niezbyt rozległym ale akumulującym się, jest to, że istnieje korelacja w populacjach pewnych gatunków miedzy poziomem infekcji a płciowością. Na przykład, wśród ślimaków w Nowej Zelandii populacje, które są zainfekowane robakami potrafiącymi je sterylizować, często rozmnażają się płciowo, podczas gdy niezainfekowane populacje pozostają bezpłciowe. Istnieją jeszcze inne dowody tego samego, korelacyjnego typu.
Teraz jednak nowy artykuł w “Science” Nadii Singh i in. (odnośnik poniżej) sugeruje, że można zobaczyć korzyści jednego aspektu seksu – rekombinacji między członkami pary chromosomów – jako reakcję poszczególnych osobników w trakcie ich życia. Zespół, używając muszek owocowych pokazał przez serię pomysłowych eksperymentów, że zainfekowane Drosophila mają wyższą proporcję rekombinowanego potomstwa niż osobniki niezainfekowane. Sugeruje to nie tylko, że pasożyty mogą być czynnikiem, który stanowił nacisk selekcyjny na płciowość u swoich gospodarzy, ale także, że gospodarze mogą w jakiś sposób „wyczuwać”, iż są zakażone i produkować bardziej zróżnicowane genetycznie potomstwo. Oczywiście, posiadanie takiego wyewoluowanego mechanizmu „wyczuwania” byłoby adaptacyjne, pozwalając na tworzenie bardziej zróżnicowanego genetycznie potomstwa – jeśli bowiem rodzic jest zainfekowany, prawdopodobnie będzie zainfekowane również potomstwo.
Tutaj są dwie muszki Drosophila melanogaster, organizm będący przedmiotem badania, uprawiające seks. Samiec, z czarnym odwłokiem, jest po prawej:

Singh i in. użyli starej techniki u Drosophila: mierzenie crossing-over (rekombinacji) między członkami jednej pary chromosomów (w ich wypadku, chromosomu 2). W ten właśnie sposób Alfred Sturtevant pokazał na początku XX wieku, że geny mieszczą się na chromosomie w określonym porządku. Był to prawdziwy geniusz. Tutaj jest jego zdjęcie mniej więcej w czasie, kiedy wykonywał ten eksperyment (proszę zwrócić uwagę na etykietkę, którą Sturtevant sam napisał):

Zespól Singh mierzył crossing-over między ebony i rough, dwoma genami na drugim chromosomie, które podlegają rekombinacji w około 20% wypadków. Stworzyli samice, które miały zmutowane geny (wpływające odpowiednio na kolor ciała i wygląd oka) na jednym chromosomie i dwa „dzikie typy” niezmutowanych genów na drugim. Ponieważ ebony i rough są genami recesywnymi , te podwójnie heterozygotyczne samice miały normalny wygląd. Kiedy jednak skrzyżuje się je z samcami homozygotycznymi dla ebony i rough, można zmierzyć ilość rekombinacji patrząc na odsetek potomstwa pokazującego tylko jedną cechę mutanta (zakreślone na diagramie poniżej, który jest wzięty z artykułu). Normalnie widzimy około 20% takiego potomstwa – w im większej odległości są geny na chromosomie, tym więcej zachodzi crossing-over i tym więcej rekombinacji widzimy.
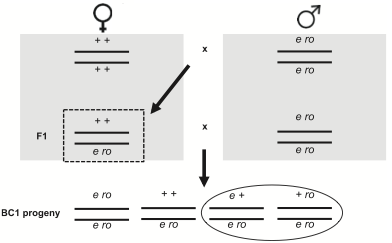
Okazuje się, że kiedy podwójnie heterozygotyczne samice są zainfekowane bakterią Serratia marcescens, po kopulacji z samcami z ebony rough, mają znacznie więcej rekombinowanego potomstwa niż zarówno niezainfekowane samice, jak te z udawanym zainfekowaniem (tj. nakłute igłą, ale bez wstrzyknięcia bakterii).
Infekcja wyraźnie podniosła stopień rekombinacji – jeden z aspektów rozmnażania płciowego – chociaż wartość p, która pokazuje prawdopodobieństwo, że te wyniki mogły być po prostu losowym wahaniem i że nie ma żadnego rzeczywistego efektu, jest trochę wyższa niż chciałbym. Wynosiła 0,03, co znaczy, że nawet gdyby nie było żadnego efektu infekcji na rekombinację, eksperymentatorzy zaobserwowaliby tej wielkości efekt w jednej z 30 prób.”Górną granicą” jest 0,05, więc biolodzy uważają wartości p poniżej tego za „istotne”. Te rezultaty są więc marginalnie istotne, ale wolałbym zobaczyć więcej eksperymentów, szczególnie takich, które pokazują, że wzrost rekombinacji obejmuje cały genom, nie zaś tylko część jednego chromosomu.
Jest możliwe z przyczyn, których nie rozumiemy, że infekcja po prostu rozłamuje chromosomy, a szczególnie chromosom drugi. Można sprawdzić także to przez zobaczenie, czy infekcja innych organizmów, na przykład małych robaków, miałaby ten sam efekt. Inny eksperyment przy użyciu zabitych bakterii, nie dał tego efektu, czyli do stworzenia bardziej zróżnicowanego potomstwa potrzebne są żywe bakterie. Co ważniejsze, eksperyment z osami (patrz poniżej) dał podobne wyniki.
Są jeszcze dwa inne wyjaśnienia wzrostu odsetka zmutowanego potomstwa. Jednym jest rekombinacja mitotyczna: crossing-over, które zachodzi nie podczas tworzenia plemnika i jajeczka (w czasie, kiedy “chromosomy siostrzane” ustawiają się w pary) ale przedtem. Albo też może być wypaczenie transmisji, czyli nie ma wzrostu w liczbie crossing-over, ale rekombinowane chromosomy w jakiś sposób zostają preferencyjnie upakowane w jajeczka, robiąc wrażenie, że nastąpił wzrost crossing-over. To nadal mogłoby – we wszystkich wypadkach – reprezentować adaptacyjną reakcję ewolucyjną, ponieważ wszystkie te trzy mechanizmy dają wyższą proporcję bardziej zróżnicowanego potomstwa. Mówimy tutaj po prostu o mechanizmie uzyskania bardziej zróżnicowanego potomstwa.
Nie będę się tutaj zagłębiał w szczegóły, ale można przetestować te alternatywne mechanizmy przez wykorzystanie dwóch cech Drosophila: samce nie wykazują crossing-over podczas tworzenia plemników (nie mamy pojęcia dlaczego tak jest!) oraz cała „normalna” rekombinacja zachodzi 4-5 dni przed złożeniem zapłodnionych jajeczek. Ergo, jeśli pojawienie się rekombinowanego potomstwa wzrasta po tym, musi to być wypaczenie transmisji. Okazuje się, że wzrost liczby rekombinowanego potomstwa istotnie pojawia się z powodu wypaczenia transmisji: rekombinowane chromosomy są preferencyjnie wkładane w jajeczka. Nie mamy pojęcia, jak to jest robione.
Wreszcie, autorzy wykonali jeszcze jeden eksperyment wstrzykując Drosophila larvae pasożytnicze osy zamiast bakterii (te osy są normalnymi pasożytami larw muszych w naturze). I, podobnie jak z bakteriami, larwy-samice, które skutecznie zwalczyły infekcję osami, dorosły i miały większy odsetek rekombinowanego potomstwa niż kontrolne, niezainfekowane larwy. Poniżej pokazana jest różnica, która jest istotniejsza (p = 0,0002) niż przy bakteriach. Odsetek rekombinowanego potomstwa jest po lewej i jest niższy w grupie kontrolnej niż w grupie zainfekowanej osami:
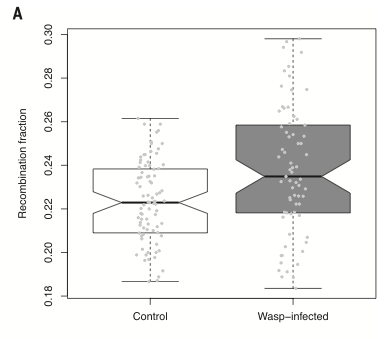
Ten wynik dokłada się więc do rosnącej sterty danych, które sugerują, że przynajmniej w niektórych wypadkach ewolucja płci związana jest z odpornością na czynniki zakaźne – tj. że seks jest odpowiedzią adaptacyjną, która odpiera infekcję przez produkowanie tak wielu różnych potomków, że niektóre z nich będą miały zestaw genów odpornych na infekcję. Lub, mówiąc inaczej, geny na rozmnażanie płciowe są korzystne, ponieważ są obecne w tych osobnikach, które mają inne geny, pozwalające im na lepsze przetrwanie infekcji. Geny płci „jadą autostopem” na genach opierających się infekcji.
Artykuł zespołu Singh rozszerza poprzednie prace, ponieważ pokazuje, że reakcja adaptacyjna może wyewoluować nie tylko na przestrzeni pokoleń, ale może być plastyczna: rekombinacja może wzmóc się podczas jednego życia po odkryciu infekcji. (Oczywiście, ta reakcja, jeśli jest prawdziwie adaptacyjna i nie jest tylko epifenomenem, musi także ewoluować przez pokolenia.)
Czy więc infekcja jest ogólną siłą selektywną, która daje płciowość w większości gatunków?
Kto wie? Płciowość jest niemal uniwersalna w organizmach wielokomórkowych, ale czy wszystkie one były poddane silnej selekcji przez czynniki zakaźne, która przezwyciężyła wysoki koszt rozmnażania płciowego? Oczywiście, kiedy już raz rozwinie się skomplikowany aparat do rozmnażania płciowego, trudno jest cofnąć się, a więc jest rodzaj inercji, która może zachować rozmnażanie płciowe, nawet jeśli nie jest już dłużej korzystne. Wbrew temu jednak jest wiele wypadków gatunków rozmnażających się płciowo, które rozmnażają się również czasami bezpłciowo. Czyli to zawrócenie ewolucji może zdarzyć się i zdarza się. Dlaczego nie dzieje się to częściej, skoro są takie korzyści z rozmnażania bezpłciowego? Czy infekcja jest tak wszechobecna i jest tak silnym naciskiem selekcyjnym? Czy też istnieją inne czynniki, które faworyzują rozmnażanie płciowe?
Tutaj jest kilka pytań, które rozszerzyłyby tę pracę, która – bez wątpienia – jest bardzo dobra.
- Czy spowodowana przez infekcję “rekombinacja” (tj. wypaczenie transmisji) u muszek jest specyficzna dla drugiego chromosomu? Byłoby miło wiedzieć, czy rekombinacja jest zwiększona w całym genomie, czego można się spodziewać, jeśli hipoteza Czerwonej Królowej jest poprawna. To jest łatwy eksperyment do przeprowadzenia.
- Czy można pokazać ewolucję wyższej rekombinacji w laboratorium? Można ustawić “klatki populacji”, każda zawierająca genetycznie zróżnicowane muszki, a potem zainfekować połowę klatek, zostawiając drugą połowę jako grupę kontrolną. Po około roku (mniej więcej 25 pokoleń) można oczekiwać przejawów ewolucji wyższej rekombinacji w zakażonych klatkach. Najlepiej byłoby kolejno zakażać muszki różnymi czynnikami zakaźnymi, żeby narzucić im nieustanną selekcję. Jeśli w pełni zaadaptują się do danego czynnika zakaźnego – który by im już nie szkodził – ustałby nacisk selekcyjny na rozmnażanie płciowe.
Na koniec powinienem dodać, że od dawna wiedzieliśmy, iż dobór sztuczny na zwiększoną rekombinację odnosi wielkie sukcesy w Drosophila. Na przykład, można wziąć rodzaj samic pokazanych powyżej i wybrać do dalszej hodowli te, które mają większy odsetek rekombinowanego potomstwa. Po kilku pokoleniach doprowadzi to do dość silnego wzrostu rekombinacji (można również selekcjonować na zmniejszenie tego wzrostu). Ponadto, można także selekcjonować na zwiększoną rekombinację między daną parą genów na jednym chromosomie, pozostawiając resztę genomu bez zmiany. Najwyraźniej istnieją geny z wariantami, które potrafią zmienić poziom rekombinacji, co oczywiście jest warunkiem koniecznym, jeśli ten aspekt płciowości wyewoluował jako adaptacja.
_____
Singh, N. D. et al. 2015. Fruit flies diversity their offspring in response to parasite infection. Science 349:747-750.
why-sex-experiments-on-fruit-flies-suggest-it-evolved-to-resist-infection
Why Evolution Is True, 16 sierpnia 2015
Tłumaczenie: Małgorzata Koraszewska

Profesor na wydziale ekologii i ewolucji University of Chicago, jego książka "Why Evolution is True" (Polskie wydanie: "Ewolucja jest faktem", Prószyński i Ska, 2009r.) została przełożona na kilkanaście języków, a przez Richarda Dawkinsa jest oceniana jako najlepsza książka o ewolucji. Jerry Coyne jest jednym z najlepszych na świecie specjalistów od specjacji, rozdzielania się gatunków. Jest wielkim miłośnikiem kotów i osobistym przyjacielem redaktor naczelnej.