Tam, gdzie są ptaki
Carl Zimmer w „New York Times” zwraca uwagę na opublikowany wczoraj w “Science” artykuł Kierena Mitchella i jego współpracowników z University of Adelaide, który zajmuje się starym problemem w biogeografii, przedstawiając nowe dane. Inny artykuł, w “Molecular Biology and Evolution” Allana Bakera i współpracowników z Royal Ontario Museum, zajmuje się tym samym problemem – i konkretnym przykładem – w podobny sposób.

Problemem jest dysjunkcja – spokrewnione gatunki znajdowane w bardzo odległych od siebie miejscach. Oto problem: w jaki sposób pokrewne gatunki znalazły się tak daleko od siebie? Szereg odpowiedzi proponowano dla zwierząt lądowych i wszystkie propozycje okazały się prawdziwe w niektórych wypadkach:
Przestrzeń pomiędzy nimi była kiedyś zamieszkała, ale stworzenia wymarły od tego czasu. Jest to dobrze znane zjawisko: na przykład, gatunek rozszerza swój zasięg w czasach przyjaznych klimatycznie. a potem ten zasięg kurczy się, gdy klimat pogarsza się, pozostawiając oddzieloną resztkę w miejscu, które nadal są przyjazne warunki. Żaba leśna (Rana sylvatica), gatunek północnoamerykański znajdowany na chłodniejszych obszarach i sporych wysokościach, ma pokrewną populację w Górach Skalistych, pozostawioną tam, kiedy zasięg przesuwał się na północ w miarę ocieplania klimatu w okresie interglacjalnym (co dla żab leśnych było pogorszeniem warunków klimatycznych).
Dysjunkcja przez wymarcie populacji na obszarze pomiędzy dwiema skrajnymi populacjami, choć stosuje się do zwierząt, które rozdziela ląd, nie może wyjaśnić dysjunkcji poprzez wodę. Doprowadziło to do popularnego kiedyś wyjaśnienia:
Most lądowy łączył kiedyś rozdzielone obszary, pozwalając organizmom na zasiedlenie ich, ale obecnie most zapadł się i jest pod wodą. Biogeografowie kreślili na oceanach hipotetyczne mosty lądowe, ale większość z nich uważa się obecnie za nieprawdopodobne z punktu widzenia geologii i niepotrzebne z punktu widzenia biogeografii. Niemniej mosty lądowe niewątpliwie istniały, a następnie znikały i organizmy przechodziły nimi, kiedy były ponad wodą. Na przykład mamuty przeszły między Azją a Ameryką w czasie niższego poziomu morza, kiedy Cieśnina Beringa stała się mostem lądowym Beringa.
Jeśli nie było połączenia lądowego, to jak organizmy przedostawały się z jednego miejsca do drugiego?
Organizmy rozprzestrzeniały się przez wodę – latając, pływając, przenosząc się na pływających przedmiotach itd. – co Darwin nazywał „przypadkowymi środkami transportu”. Dla mas lądowych, które nigdy nie były połączone, był to jedyny sposób na pojawienie się dysjunkcji i zaobserwowano wiele przykładów rozprzestrzeniania się przez wodę, najczęściej przez lot, ale bezpośrednio zaobserwowano także rzadsze sposoby (np. jaszczurki przenoszące się z wyspy na wyspę w Indiach Zachodnich, żółw olbrzymi Aldabra wychodzący na ląd w Tanzanii).
I wreszcie istnieje możliwość, która stała się popularna po odkryciu ruchu płyt tektonicznych w geologii:
Dysjunkcja była spowodowana rozdzieleniem się pierwotnie ciągłej masy lądowej na dwie lub więcej części, tak że geograficzna dystrybucja organizmów rozrzuconych po całej masie lądowej została także rozdzielona. Innymi słowy, organizmy nie ruszyły się z miejsca – zrobiły to kontynenty. Kiedy geologia pokazała rzeczywistość dryfu kontynentalnego, stało się jasne, że dysjunkcja może zdarzyć się nie z powodu przenoszenia się zwierząt (chociaż pierwotnie musiały rozprzestrzenić się po ciągłym lądzie pierwotnym), ale przez przesuwanie się połaci ziemi. Żeby więc użyć przykładu skamielin, podobny do ssaków gad Cynognathus z triasu został znaleziony zarówno w Afryce, jak w Ameryce Południowej. To korpulentne zwierze o długości metra nie było dobrym kandydatem na podróże przez oceany; kiedy jednak zrozumiano, że w owym czasie te dwa kontynenty były połączone, problem zniknął; dryf kontynentalny wyjaśnił dysjunkcję.
Problemem badanym zarówno przez Mitchella, jak Bakera jest dystrybucja i stosunki pokrewieństwa strusiowatych – grupy dużych, nielotnych ptaków, która obejmuje strusie, nandu, kazuary, kiwi, emu i wymarłe niedawno moa oraz mamutaki.
Wszystkie te ptaki znajdują się głównie na kontynentach południowych, które były kiedyś połączone w jeden super-kontynent Gondowanę. Oto ich obecna dystrybucja:
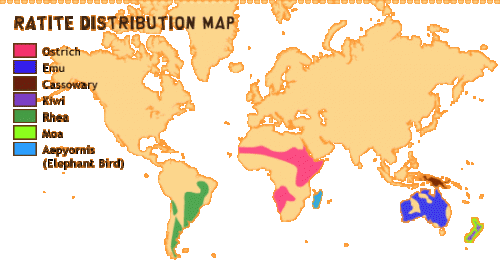
Od dawna było wiadomo, że strusiowate są wzajem ze sobą spokrewnione, jak również są spokrewnione z kusaczami, marnie latającymi ptakami z Ameryki Południowej. Jak te różne nielotne formy znalazły się w swoich obecnych miejscach zamieszkania było przez długi czas zagadką. Rozprzestrzenianie się ponad wodą wydawało się wykluczone (przecież są to ptaki nielotne) więc może istniały kiedyś na kontynentach północnych, ale wymarły tam. Po pojawieniu się informacji o ruchu płyt tektonicznych ulubioną hipotezą było, że przodek strusiowatych był szeroko rozprzestrzeniony na Gondowanie i, kiedy super-kontynent rozłamał się w krecie, nielotne ptaki, każdy na swoim kawałku lądu, poszły swoimi drogami i wyewoluowały w dzisiejsze gatunki. Orędownikiem tego poglądu był Joel Cracraft, pracujący obecnie w American Museum.
Ten pogląd jednak spotkał się z krytyką i różni autorzy mówili, że ptaki jednak rozprzestrzeniały się ponad wodą (patrz na przykład to streszczenie wykładu Mike’a Dickisona z 2008 r.). Obie nowe prace, Mitchella i Bakera, popierają pogląd rozprzestrzenienia się ponad wodą. Baker, używając technik badania prastarego DNA do zbadania DNA wymarłych moa, potwierdza wcześniejsze prace własne i jego współpracowników pokazujące, że najbliższymi krewnymi moa są latające kusacze, pokazując w ten sposób, że nielotność wyewoluowała więcej niż jeden raz wśród strusiowatych. Mitchel także używa prastarego DNA i pokazuje, że najbliższymi krewnymi mamutaków z Madagaskaru są kiwi z Nowej Zelandii, ponadto ocenia czas rozejścia się na 50 milionów lat temu – długo po tym, jak Gondwana rozpadła się do tego stopnia, że szerokie morze oddzielało Madagaskar od Nowej Zelandii, co sugeruje, że wspólny przodek tych dwóch ptaków latał.
Obie grupy badaczy wyciągają wniosek, że większość lub wszystkie wzory dystrybucji strusiowate zawdzięczają migracjom ponad wodą swoich latających przodków, co zdarzyło się długo po rozłamanie się Gondwany na jej nowoczesne części i że nielotność wyewoluowała zbieżnie u strusiowatych. Według „Timesa”, Cracraft jeszcze nie jest całkowicie przekonany, ale sądzi, że dalsze badania wkrótce pokażą pełniejszą historię ewolucyjną strusiowatych.
______________________________________________________________
Baker, A.J., Haddrath, O., McPherson, J.D., and A. Cloutier.2014. Genomic support for a moa-tinamou clade and adaptive morphological convergence in flightless ratites. Molecular Biology and Evolution in press.
Cracraft, J. 1973. Phylogeny and evolution of the ratite birds. Ibis 116:494-521.
Mitchell, K.J., Llamas, B. Soubrier, J, Rawlence, N.J., Worthy, T.H., Wood, J., Lee, M.S.Y., and A. Cooper. 2014. Ancient DNA reveals elephant birds and kiwi are sister taxa and clarifies ratite bird evolution. Science 344:898-900.
Why Evolution Is True, 24 maja 2014
Tłumaczenie: Małgorzata Koraszewska

Greg Mayer
Zastępca kuratora działu gadów i płazów w muzeum zoologicznym Uniwersytetu Wisconsin. Doktorat z biologii ewolucyjnej obronił na Harvardzie.