Nie mniej niż dziesięcioro czytelników zwróciło mi uwagę na
nowy artykuł w „Nature Communications” Anny M. Kearns i in. (odnośnik na dole, pdf
tutaj), pokazujący podobno “odwrócenie specjacji” u kruków. Artykuł zyskał wiele publicznej uwagi, bo autorzy twierdzą, że pokazują jak dwa różne gatunki kruków zlały się ponownie w jeden gatunek. To zaś podekscytowało ludzi, ponieważ a.) nie sądzą, by to zdarzyło się kiedykolwiek wcześniej; i b.) pokazuje to, że specjacja nie jest prostym, rozgałęziającym się drzewem, jakie przedstawił Darwin i że niektóre gałęzie mogą zrosnąć się z powrotem w jedną, nową gałąź.
Ale, jak piszą autorzy, widzieliśmy już wcześniej fuzje gatunków. Często jest to związane z antropogenicznymi zmianami habitatu, takimi jak zmiana klimatu, lokalnej ekologii, lub introdukcji drapieżników (patrz streszczenie artykułu tutaj). Możemy to obecnie zobaczyć z niedźwiedziem polarnym i niedźwiedziem grizli. Izolowane poprzednio przez ekologiczne preferencje i adaptację (oparta na genach „reprodukcyjna bariera izolacyjna”) ich ekologiczne rozdzielenie może zniknąć wraz ze zmianą klimatu. Kiedy niedźwiedź grizli przesunie się na północ wraz z ociepleniem klimatu, wejdzie na terytorium niedźwiedzia polarnego i te dwa gatunki mogą tworzyć hybrydy i robiły to wielokrotnie w naturze. Jeśli hybrydy są płodne (nie mogę znaleźć danych na ten temat), te dwa gatunki mogą zlać się w jeden.
Są jednak inne przypadki fuzji gatunków, za które ludzie nie są odpowiedzialni. W końcu zmiany ekologiczne, zmiana klimatu i introdukcja drapieżników mogą zdarzyć się bez interwencji ludzi. Problem ze znalezieniem hybrydowych gatunków w naturze polega na tym, że niełatwo wykryć, iż są wynikiem hybrydyzacji, jeśli oba gatunki rodzicielskie wymarły. (Hybrydową naturę gatunku wykrywa się zazwyczaj po tym, że są genetyczną mozaiką rodzicielskich gatunków, a jeśli rodziców nie ma, to trudno to zaobserwować.) Mamy jednak wiele przykładów „hybrydowych gatunków”, które nie zastąpiły gatunków rodzicielskich, włącznie z wieloma allopoliploidalnymi roślinami, jak również diploidalnymi hybrydowymi gatunkami motyli i słoneczników. Ponadto gatunki wymieniają geny częściej niż myśleliśmy dawniej i ten „horyzontalny transfer genów” może zabałaganić filogenetyki.
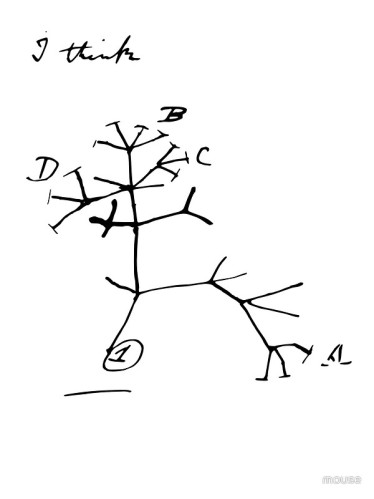
Nic z tego jednak nie obala uogólnienia, że gatunki niemal zawsze powstają z geograficznie izolowanych populacji, które różnicują się genetycznie do punktu, kiedy już nie mogą wymieniać genów, bo wyewoluowały bariery do wymiany genów takie jak bezpłodność hybryd, ekologiczne preferencje i adaptacje, wybieranie partnerów i tak dalej. Poza roślinami hybrydowa specjacja jest rzadkim wyjątkiem, nie zaś regułą, i „rozgałęziające się drzewo” życia Darwina, naszkicowane w jego notatniku, nadal jest dobrym opisem życia:
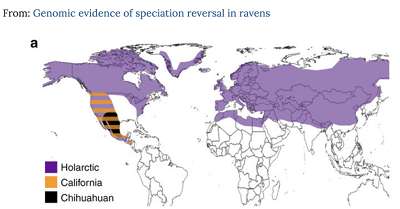
Co więc znaleźli Kearns i in.? Odkryli, że dwie stare linie rodowe kruków zwyczajnych (Corvus corax), jeden szeroko rozprzestrzeniony (“holarktyczny”) i jeden z zachodniego wybrzeża Ameryki Północnej (“kalifornijski”), rozeszły się, prawdopodobnie po geograficznej izolacji, około 1,5 miliona lat temu, ale potem nie stały się istniejącymi gatunkami, ponieważ zaczęły wymieniać geny – wielokrotnie. (Geograficzna izolacja mogła wynikać z izolowania wspólnego przodka w ostojach w okresach zlodowacenia.) Teraz więc w zachodnich USA i w Meksyku znajduje się kruki, które wyglądają tak samo, jak inne kruki na całym świecie, ale mają geny pradawnej linii rodowej, która musiała oddzielić się od „holoarktycznych” kruków bardzo dawno temu. Można to wykryć, ponieważ na tym terenie znajdują się ptaki, które mają dwie kopie genów, które rozeszły się w pradawnych czasach – są znacznie odleglejsze od siebie niż normalna zmienność w ramach holarktycznych kruków.
Obecnie postaci holarktyczna i kalifornijska nie są różnymi gatunkami, ponieważ hybrydyzują z łatwością na zachodzie Ameryki Północnej i nie ma dwóch różnych „typów”. Można zobaczyć je na poniższej mapie: holarktyczne są liliowe, kalifornijskie są pomarańczowe, a hybrydy – mając geny obu – są w paski fioletowo pomarańczowe (nie znajduje się samych pomarańczowych; ich geny po prostu zlały się z genami holarktycznych kruków).
Sytuacja jest skomplikowana, ponieważ jest tam jeszcze jeden gatunek kruka, który jest „sympatryczny” (żyje na tym samym obszarze, co) zarówno holarktyczne, jak kalifornijskie kruki, ale pozostaje odrębny: kruk meksykański, zamieszkujący czarne obszary poniżej. Jest uważany za inny gatunek (Corvus cryptoleucus), ponieważ nie ma dowodów, że hybrydyzuje z jakąkolwiek inną grupą; wydaje się w pełni izolowany reprodukcyjnie od pozostałych kruków.
Dystrybucja tych dwóch linii rodowych (i dwóch gatunków: zwyczajnego i meksykańskiego) z artykułu:
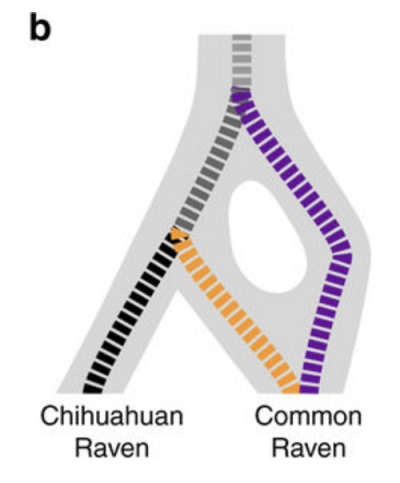
Sekwencjonując DNA autorzy odkryli, że kruk meksykański jest bliżej spokrewniony z linią kalifornijską kruka zwyczajnego niż z linią holarktyczną i najwyraźniej oderwał się od kalifornijskiej później niż rozejście się linii meksykańskiej i holarktycznej. Gałąź meksykańska oderwała się od linii kalifornijskiej między 0,6 a 1,5 miliona lat temu.
To, co mamy, pokazane jest na diagramie poniżej: nierozgałęziające się drzewo rodzinne. Odzwierciedla ono prastare rozejście się linii, które dało holarktyczną gałąź kruka zwyczajnego z jednej strony i kruka meksykańskiego + kalifornijska linię z drugiej. Ta gałąź rozdzieliła się później znowu, dając gatunek kruka meksykańskiego i linię rodową kruka zwyczajnego, która zlała się z powrotem z linia holarktyczną. To powinno być jasne na diagramie:
Rezultat: Wielkim pytaniem i powodem, dla którego ten artykuł zyskał tak wiele uwagi, jest to, że pokazał „odwrócenie specjacji”. To jest, autorzy zakładają, że linia kalifornijska była kiedyś gatunkiem kruków odrębnym od holarktycznych kruków zwyczajnych, ale zlała się z nim później. W jakiś sposób bariery reprodukcyjne (genetyczne), które trzymały je osobno, stały się nieskuteczne.
Oczywiście zakłada to, że kiedyś istniały bariery reprodukcyjne między liniami kalifornijską i holarktyczną. Ale nie mamy na to żadnych dowodów! Fakt, że tak łatwo zlały się, przemawia przeciwko znaczącym barierom reprodukcyjnym, choć mogły istnieć jakieś bariery ekologiczne. Autorzy po prostu zakładają, że ponieważ kruki meksykańskie stały się gatunkiem w krótszym czasie niż rozejście się holarktycznych-kalifornijskich, to rozejście się musi TAKŻE dotyczyć pełnej specjacji.
Czas rozejścia się jednak mówi nam bardzo mało o specjacji. Kluczowe jest to, czy linia rodowa ewoluuje bariery reprodukcyjne jedna od drugiej, nie zaś jak długo były rozdzielone. A te bariery reprodukcyjne są prawdopodobnie produktem ubocznym doboru, który może być silny lub słaby. Dlatego, kiedy ocenia się, czy dwie populacje są biologicznymi gatunkami, ewolucjoniści wolą używać wskaźników izolacji reprodukcyjnej (obserwacje braku kojarzenia się, żadnych dowodów domieszki genetycznej) niż czasu rozejścia się. Być może linie holarktyczna i kalifornijska po prostu nie rozeszły się genetycznie w wystarczającym stopniu, by dać jako produkt uboczny bariery reprodukcyjne. To stało się z Homo sapiens: nasze geograficznie izolowane populacje w Polinezji, Australii i Nowym Świecie nie były rozdzielone wystarczająco długo od reszty gatunku, by stać się nowym gatunkiem ludzi. Teraz, kiedy mamy transport i migracje, jesteśmy w trakcie procesu powolnego zlewania się w jedną dużą pulę genową, która może nigdy nie być w pełni zmieszana, ale będzie zmieszana w wystarczającym stopniu, byśmy mogli krzyżować się z ludźmi z każdego innego miejsca.
Autorzy widzą ten problem, ale – być może autorzy zafascynowani koncepcją „odwrócenia specjacji” (w odróżnieniu od mniej fascynującego „odwrócenia linii rodowej”), przedstawiają wadliwy „argument czasu” na rzecz specjacji (moje podkreślenia):
Tak więc nie jest całkowicie oczywiste, czy powinniśmy nazywać tę sytuację z krukami “odwróceniem specjacji”, czy uważać to za wypadek “fuzji starodawnych linii rodowych”. Kontrastuje to z większością innych przykładów odwrócenia specjacji, gdzie istnieją mocne dowody na siłę i naturę izolacji reprodukcyjnej przed odwróceniem specjacji mimo bardzo płytkiego rozejścia się między liniami rodowymi (np. cierniki). Dwie linie dowodowe sugerują, że linie kalifornijska i holoarktyczna mogły być izolowane reprodukcyjnie przed ponownym kontaktem i zlaniem się linii. Pierwsza, czas rozejścia się linii holoarktycznej i przodków linii kalifornijskiej i meksykańskiej między 0,9 a 2 miliony lat temu zbliża się do granicy, przy której większość taksonów ptaków (szczególnie tych z północnej półkuli) wyewoluowało izolację reprodukcyjną (~2 miliony lat). Druga, cechy historii życia, jak również mtDNA, intron, SNP i analiza ENM popierają [tezę o] izolacji reprodukcyjnej między krukami meksykańskimi a krukami zwyczajnymi mimo bliższego w czasie rozejścia się między krukami meksykańskimi a linią kalifornijską 0,6-1,5 miliona lat temu. Pokazuje to, że kruki mogą rozwinąć izolację reprodukcyjną i zachowywać mocne granice gatunkowe po późniejszym rozejściu się niż ta między liniami kalifornijską i holarktyczną. Twierdzimy, że nasze dane stanowią najsilniejsze możliwe poparcie dla wniosku o odwróceniu specjacji w sytuacji, kiedy nie można zmierzyć starodawnej izolacji reprodukcyjnej przed fuzją.
Nie przekonuje mnie żadna z tych linii dowodowych, że linie holarktyczna i kalifornijska były odrębnymi gatunkami. Czas specjacji może wahać się między taksonami i jeśli dobór jest słaby, linie mogą nie dochodzić do pełnych gatunków przez bardzo długi czas. Powoływanie się na dane o innych gatunkach nie rozstrzyga kwestii w tym konkretnym przypadku. Druga linia dowodów nie jest właściwie niezależna: autorzy mówią po prostu, że ponieważ krukowi meksykańskiemu specjacja zabrała mniej czasu niż rozejście się między liniami holarktyczną i kalifornijską, te dwie linie muszą także być gatunkami. To nie jest przekonujące. „Czas specjacji” bardzo się różni, nawet wewnątrz grupy, bo zależy od różnych czynników, które obejmują siłę doboru, stopień geograficznej izolacji i tak dalej.
Tutaj widzimy więc zlanie się linii, a nie zlanie się gatunków. Autorzy piszą o tym problemie (wytłuszczony tekst powyżej), ale próbują usunąć problem. Szkoda, że dziennikarze piszący o tym wiedzą tak mało o specjacji.
h/t: Michael, Jószef, Tom
___________
Kearns, A. M., M. Restani, I. Szabo, A. Schrøder-Nielsen, J. A. Kim, H. M. Richardson, J. M. Marzluff, R. C. Fleischer, A. Johnsen, and K. E. Omland. 2018. Genomic evidence of speciation reversal in ravens. Nature Communications 9:906.
”Reverse speciation” (fusion of species) in ravens
Why Evolution Is True, 7 marca 2018
Tłumaczenie: Małgorzata Koraszewska
Jerry A. Coyne
Profesor (emeritus) na wydziale ekologii i ewolucji University of Chicago, jego książka "Why Evolution is True" (Polskie wydanie: "Ewolucja jest faktem", Prószyński i Ska, 2009r.) została przełożona na kilkanaście języków, a przez Richarda Dawkinsa jest oceniana jako najlepsza książka o ewolucji. Jerry Coyne jest jednym z najlepszych na świecie specjalistów od specjacji, rozdzielania się gatunków. Jest wielkim miłośnikiem kotów i osobistym przyjacielem redaktor naczelnej.